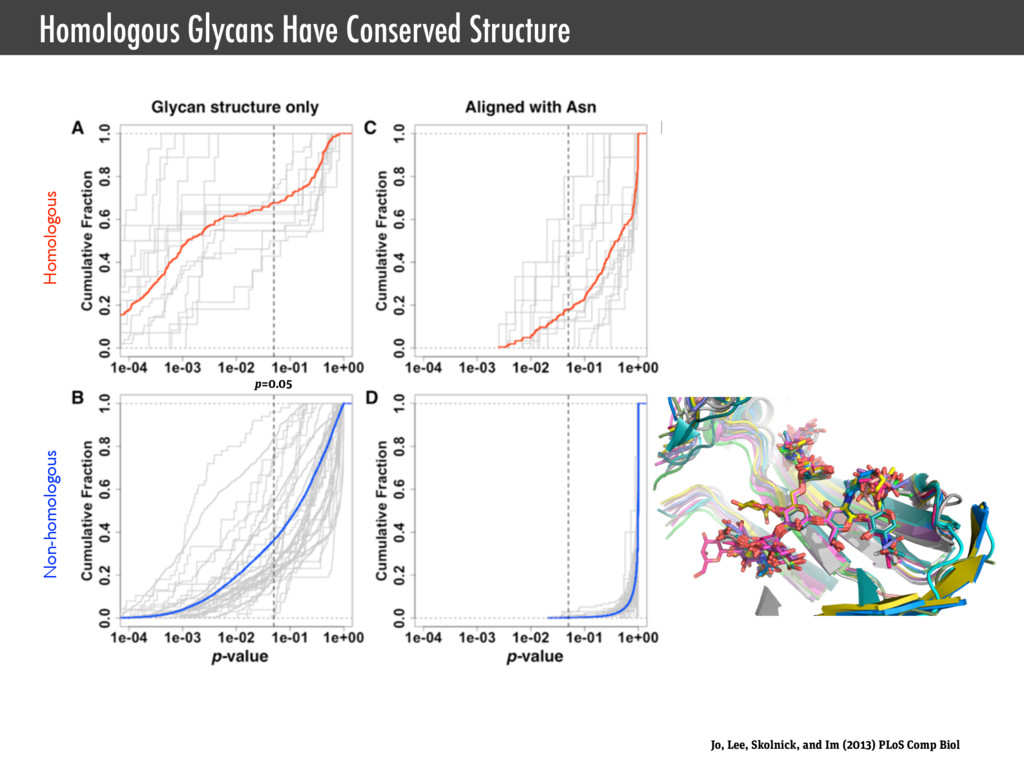
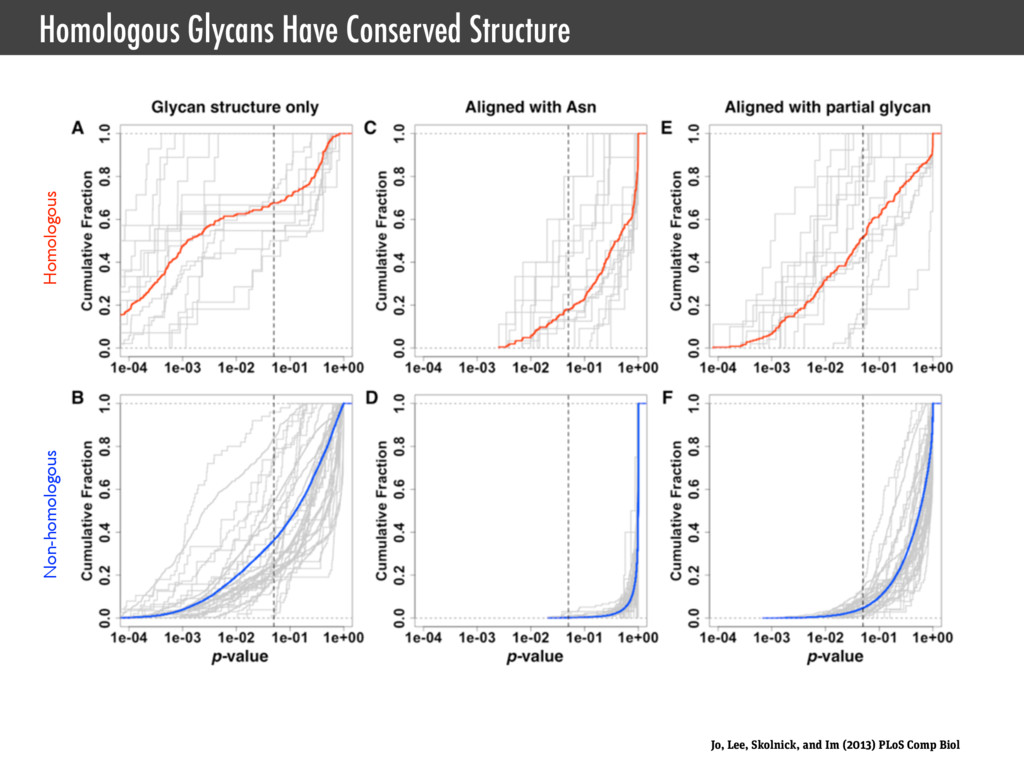
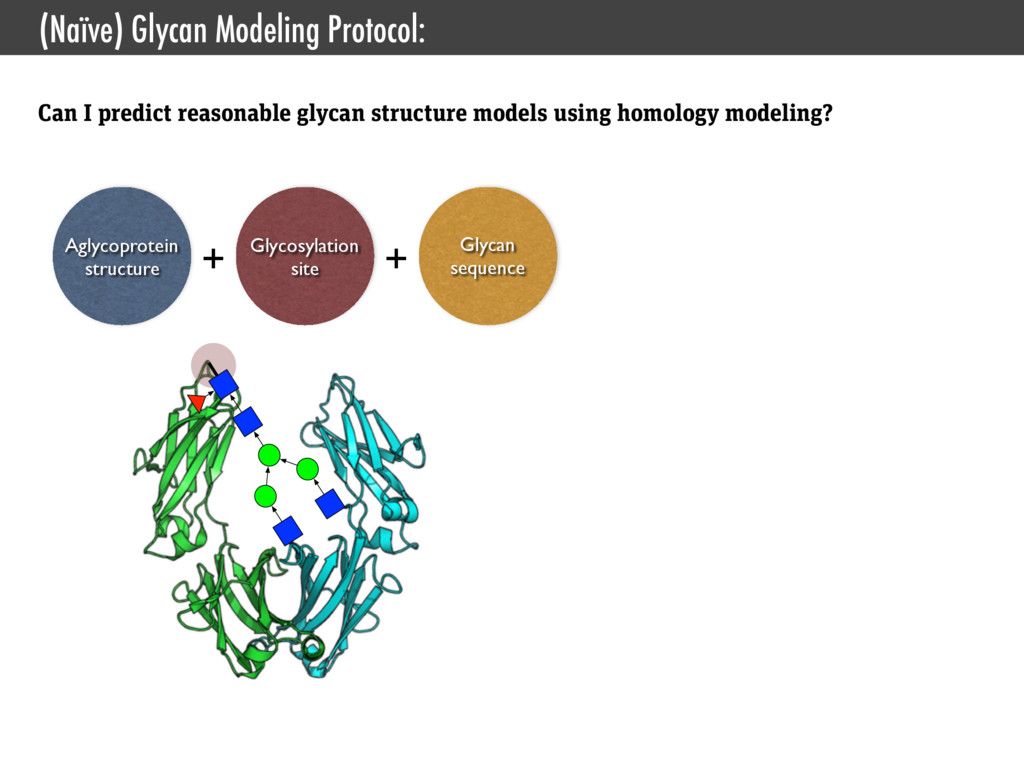
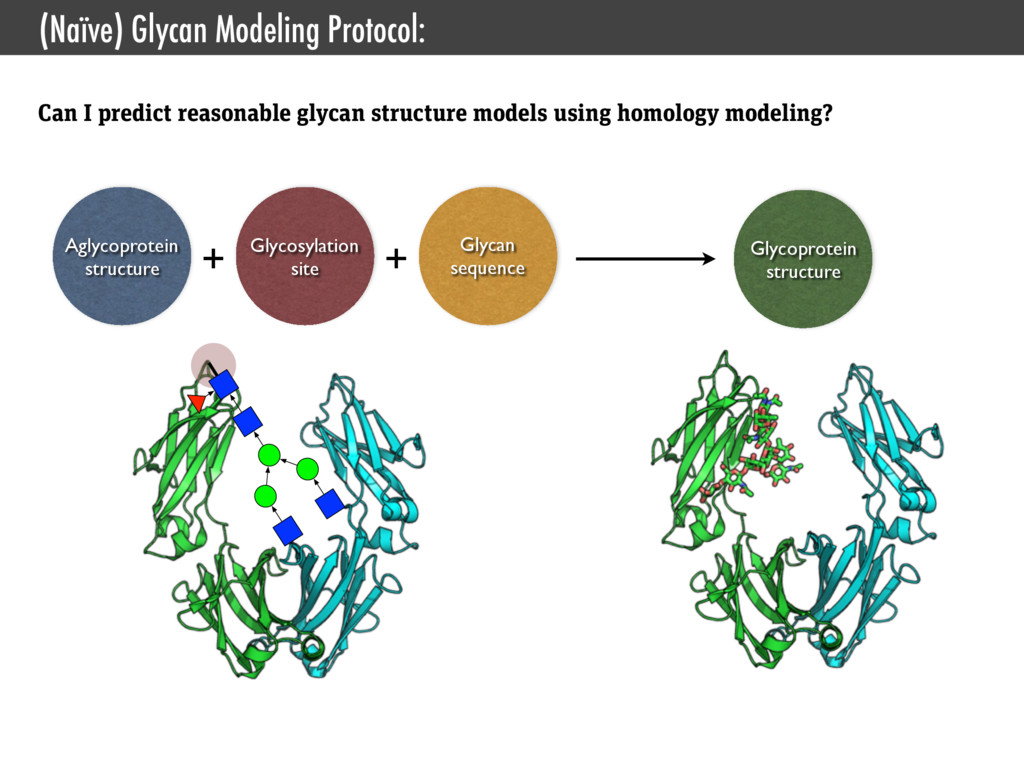
Interface for CHARMM • Jo, Kim, Iyer, and Im. (2008) CHARMM-GUI: a web-based graphical user interface for CHARMM., J. Comput. Chem. • Jo, Kim, and Im. (2007) Automated builder and database of protein/membrane complexes for molecular dynamics simulations., PLoS ONE • Jo, Vargyas, Vasko-Szedlar, Roux, and Im. (2008) PBEQ-Solver for online visualization of electrostatic potential of biomolecules., Nucleic Acids Res. • Jo, Lim, Klauda, and Im. (2009) CHARMM-GUI Membrane Builder for mixed bilayers and its application to yeast membranes., Biophys. J. • Lee, Jo, Rui, Egwolf, Roux, Pastor, and Im. (2012) Web interface for brownian dynamics simulation of ion transport and its applications to beta-barrel pores., J. Comput. Chem. • Jo, Jiang, Lee, Roux, and Im. (2013) CHARMM-GUI Ligand Binder for Absolute Binding Free Energy Calculations and Its Application., J. Chem. Inf. Model. • Cheng, Jo, Lee, Klauda, and Im. (2013) CHARMM-GUI Micelle Builder for Pure/Mixed Micelle and Protein/Micelle Complex Systems., J. Chem. Inf. Model. SSNMR Ensemble Dynamics • Jo and Im. (2011) Transmembrane helix orientation and dynamics: insights from ensemble dynamics with solid-state NMR observables., Biophys. J. • Kim, Jo, and Im (2011) Solid-State NMR Ensemble Dynamics as a Mediator between Experiment and Simulation., Biophys. J. • Im, Jo, and Kim. (2012) An ensemble dynamics approach to decipher solid-state NMR observables of membrane proteins., Biochim. Biophys. Acta Glycan Structure Modeling • Jo, Song, Desaire, Mackerell, and Im. (2011) Glycan Reader: automated sugar identification and simulation preparation for carbohydrates and glycoproteins., J. Comput. Chem. • Jo and Im. (2012) Glycan fragment database: a database of PDB-based glycan 3D structures., Nucleic Acids Res. • Jo, Lee, Skolnick, and Im. (2013) Restricted N-glycan Conformational Space in the PDB and Its Implication in Glycan Structure Modeling. PLoS Comp. Biol. • Wu, Engström, Jo, Stuhlsatz, Yeom, Klauda, Widmalm, and Im. (2013) Molecular Dynamics and NMR Spectroscopy Studies of E. coli Lipopolysaccharide Structure and Dynamics. Biophys. J. Type III Secrection System • Zhong, Lefebre, Kaur, McDowell, Gdowski, Jo, Wang, Benedict, Lea, Galan, and De Guzman. (2012) The Salmonella Type III Secretion System Inner Rod Protein PrgJ Is Partially Folded., J. Biol. Chem. Cholesterol Orientation in Lipid Bilayer • Jo, Rui, Lim, Klauda, and Im. (2010) Cholesterol flip-flop: insights from free energy simulation studies., J. Phys. Chem. B
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}