e.g. for K+ - 4 nm (cell membrane width) takes 4 nanoseconds - 1 cm takes 7 hours 1 is %me, is mean distance travelled, and is the diffusion coefficient of the substance
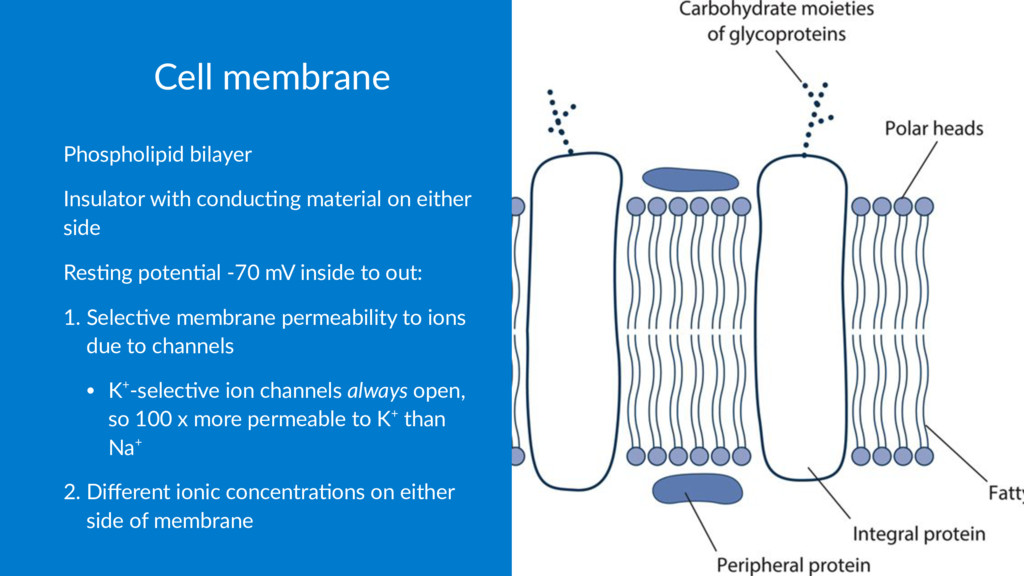
side Res$ng poten$al -70 mV inside to out: 1. Selec've membrane permeability to ions due to channels • K+-selec've ion channels always open, so 100 x more permeable to K+ than Na+ 2. Different ionic concentra'ons on either side of membrane
• Passive diffusion of K+ down its concentra*on gradient (150mM -> 5mM) out of the cell 2. Na+/K+–ATPase • 3 Na+ out, 2 K+ in • Net movement of posi*ve charge out of the cell • Only directly contributes -4mV to the res*ng poten*al • However it establishes the chemical gradient of K+ and Na+ Na+ itself contributes li1le to the res3ng poten3al as the membrane is impermeable to it.
produce over a permeable membrane 2 Simplifies at 18ºC for monovalent ions to: 2 is the equilibrium poten1al for a specific ion, the gas constant (8.314 J.K-1.mol-1), the temperature (K), Faraday's constant (96,500 C.mol-1), and the ionic valency (e.g. +1 for Na+)
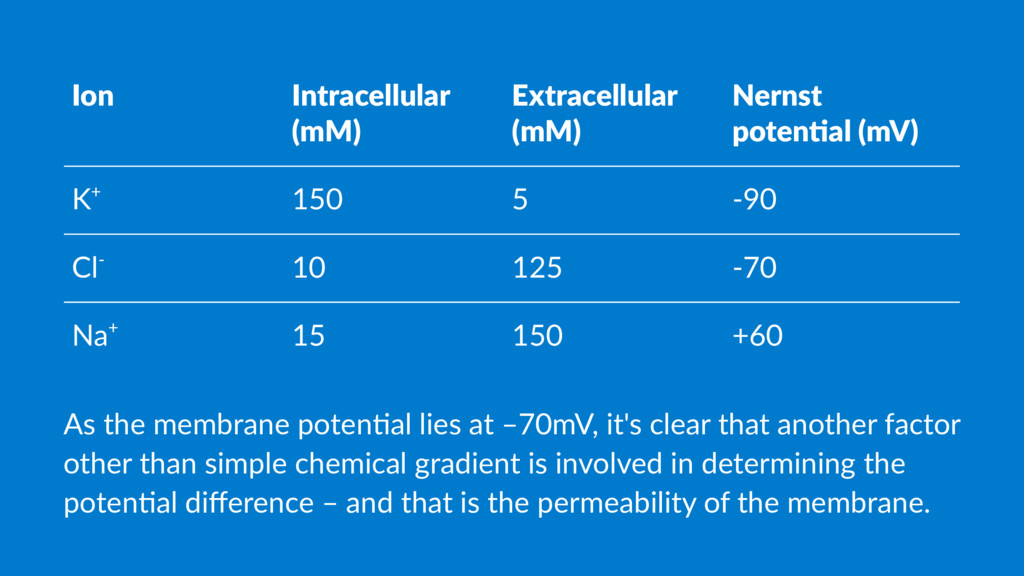
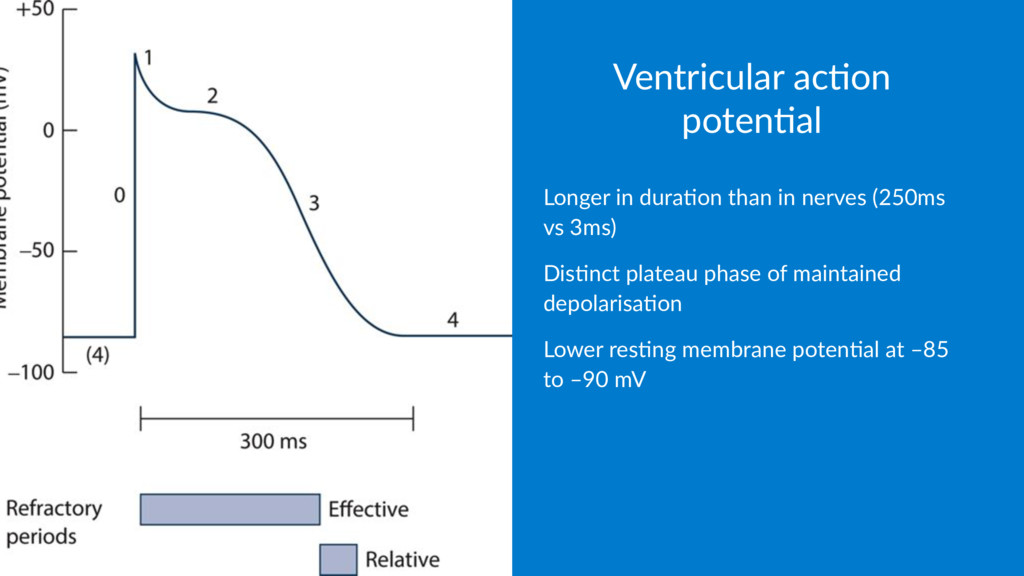
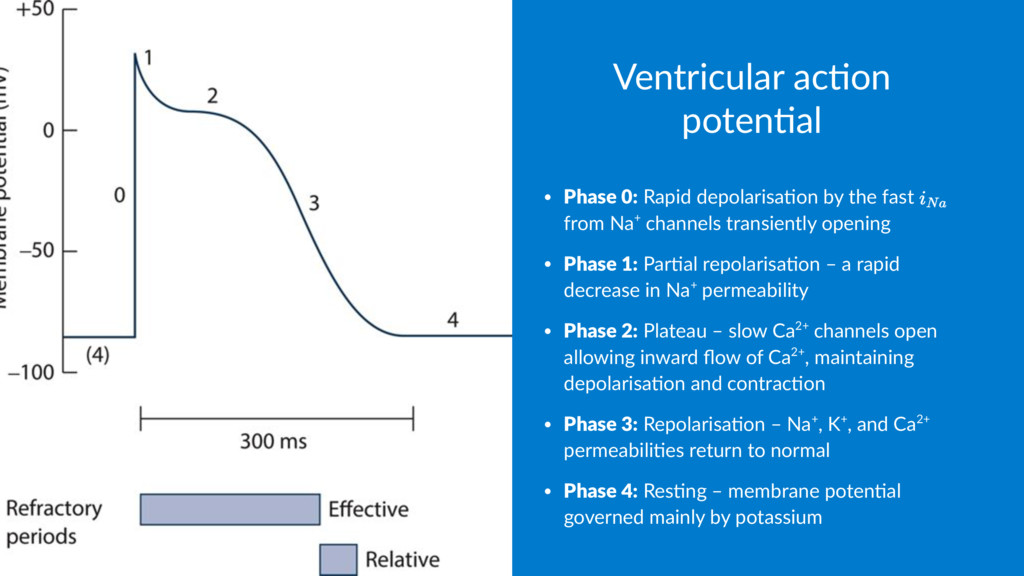
5 -90 Cl- 10 125 -70 Na+ 15 150 +60 As the membrane poten.al lies at –70mV, it's clear that another factor other than simple chemical gradient is involved in determining the poten.al difference – and that is the permeability of the membrane.
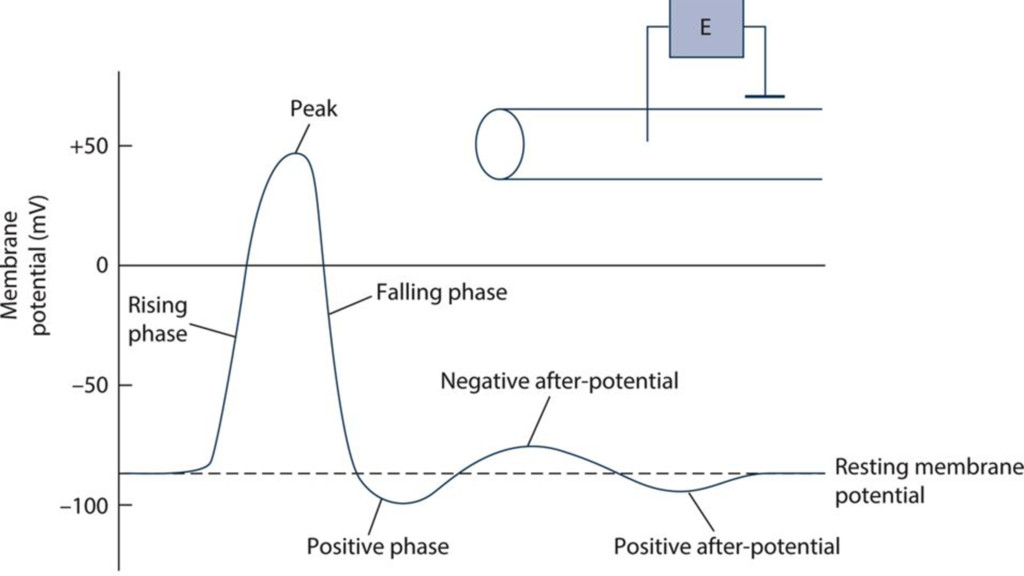
membrane polarity "All or none" response Generated by voltage–gated Na+ channels (blocked by tetrodotoxin ! and local anaesthe9cs) Threshold s*mulus: ~15 mV Dura%on: 2–3 ms
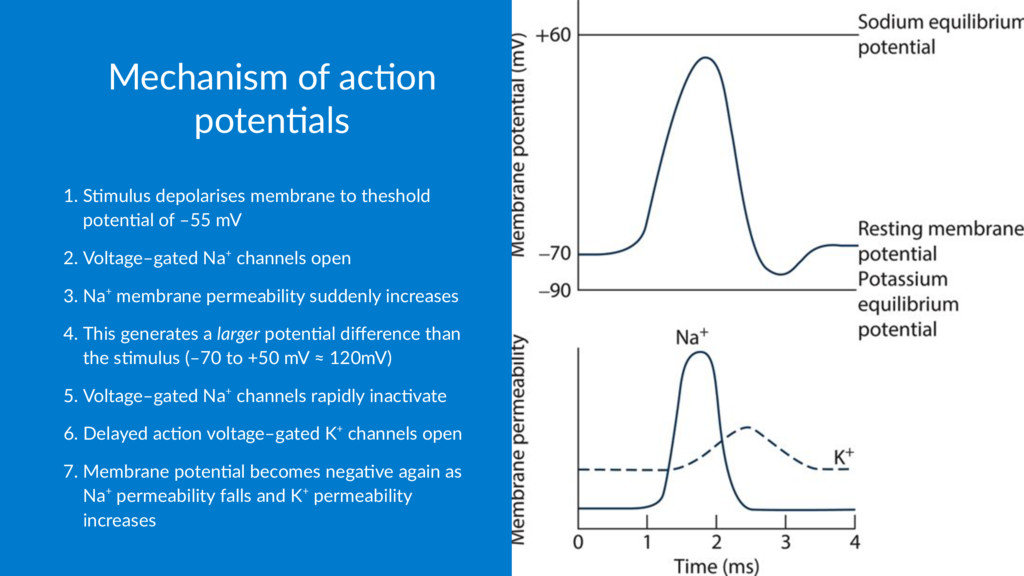
of depolarisa.on to the adjacent area of membrane via local currents 2. Regenera.on through the ac.on poten.al as the resultant poten.al difference is amplified back to full size Both posi%ve and nega%ve feedback loops are involved: • "All or none" effect is due to the posi3ve feedback loop between membrane depolarisa3on and Na+ permeability • Delayed ac3on voltage–gated K+ channels produce nega3ve feedback, returning membrane to its res3ng poten3al Only a small number of ions flow during channel opening – the change in membrane permeability is the most significant effect on poten;al difference, not the number of ions flowing
s8ll open and cell is completely inexcitable • Rela8ve refractory period: • 10–15 ms when a further ac8on poten8al can only be triggered by greater than normal s8muli
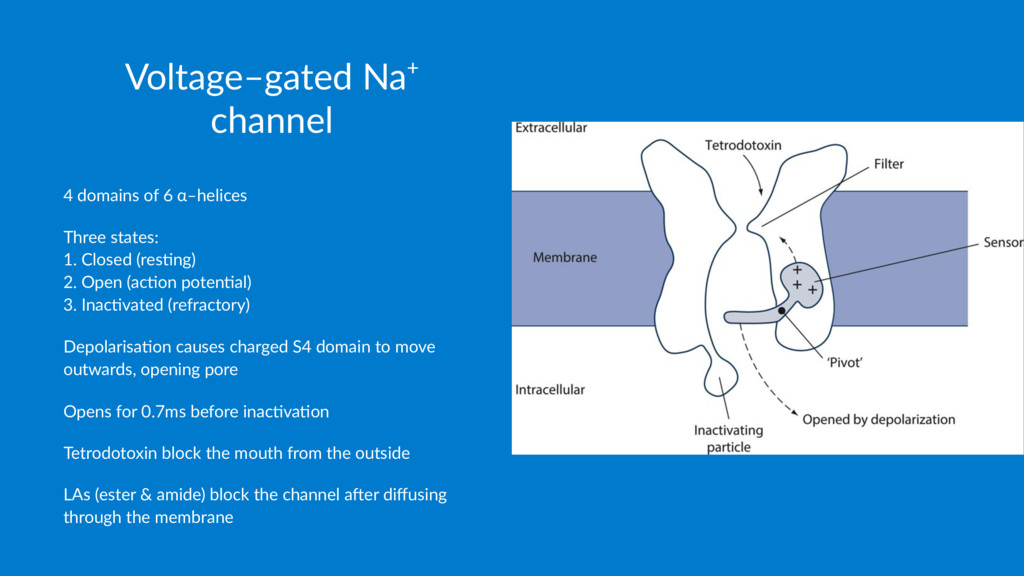
1. Closed (res1ng) 2. Open (ac1on poten1al) 3. Inac1vated (refractory) Depolarisa*on causes charged S4 domain to move outwards, opening pore Opens for 0.7ms before inac2va2on Tetrodotoxin block the mouth from the outside LAs (ester & amide) block the channel a6er diffusing through the membrane
of propoga#on than passive conduc#on due to the latency between threshold poten#al and full size depolarisa#on Cell membranes behave as capacitors as they are thin and hold charge well Myelin increases the resistance and reduces the capacitance of the membrane, so ac6on poten6als can only occur at nodes of Ranvier Between the nodes, local currents cause depolarisa3on of the next node (passive = faster) Myelin conserves energy due to the reduced ion flux requiring less energy to restore res5ng ionic concentra5ons
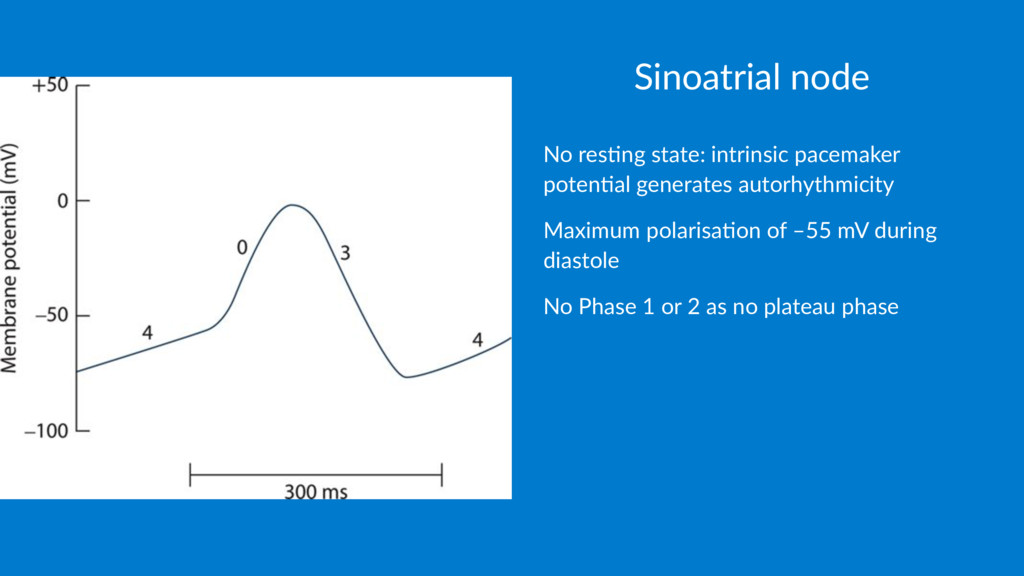
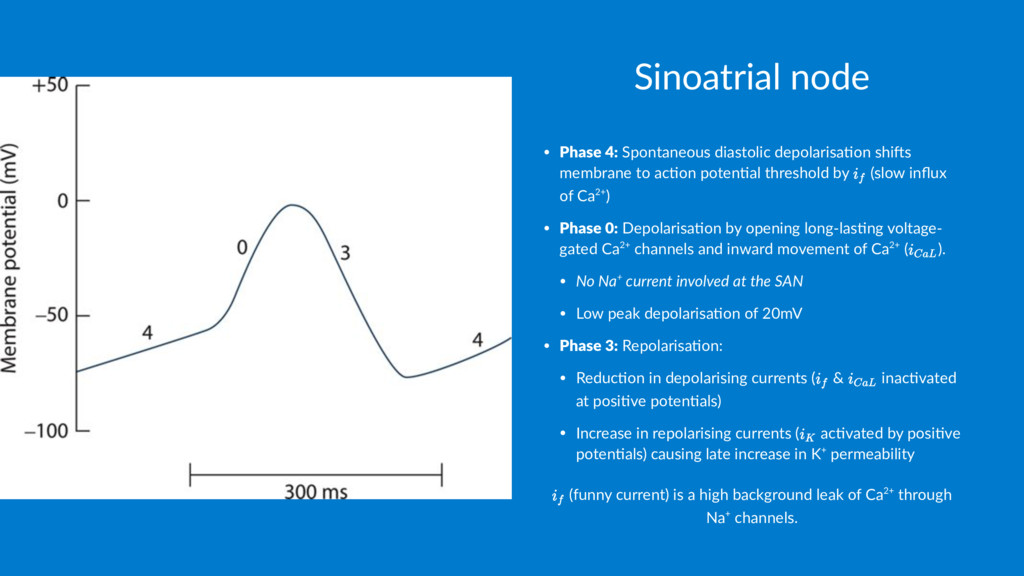
to ac1on poten1al threshold by (slow influx of Ca2+) • Phase 0: Depolarisa1on by opening long-las1ng voltage- gated Ca2+ channels and inward movement of Ca2+ ( ). • No Na+ current involved at the SAN • Low peak depolarisa1on of 20mV • Phase 3: Repolarisa1on: • Reduc1on in depolarising currents ( & inac1vated at posi1ve poten1als) • Increase in repolarising currents ( ac1vated by posi1ve poten1als) causing late increase in K+ permeability (funny current) is a high background leak of Ca2+ through Na+ channels.
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}