This talk was given by Professor Neocles Leontis at the European Bioinformatics on April 21, 2015.
Abstract:
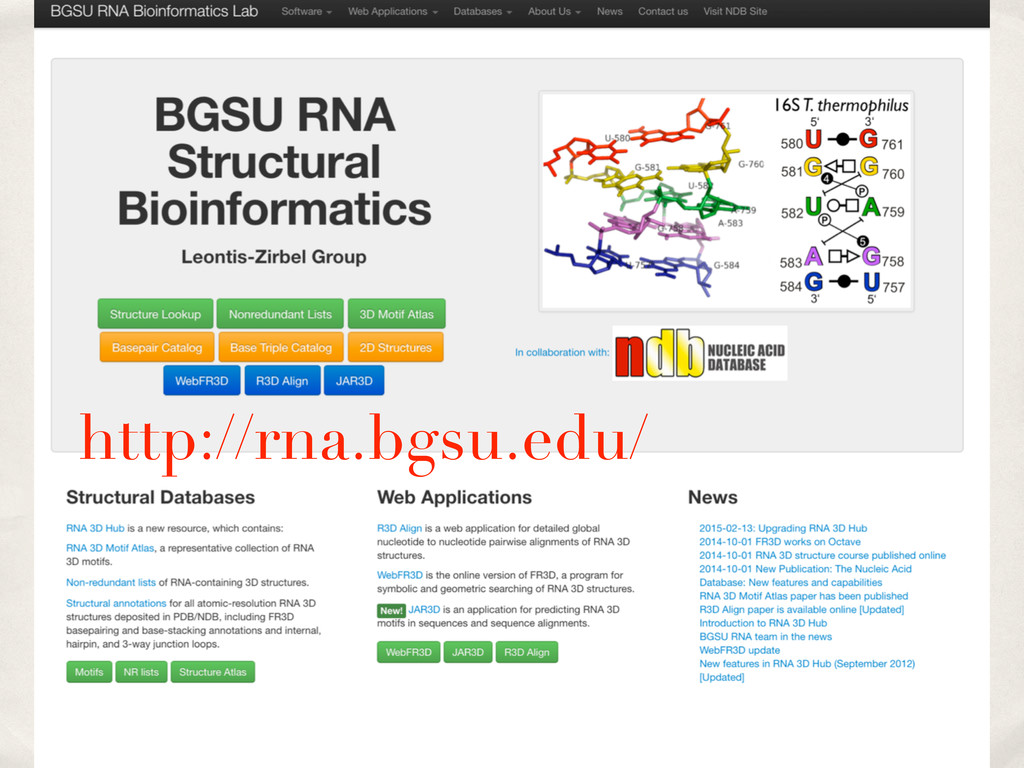
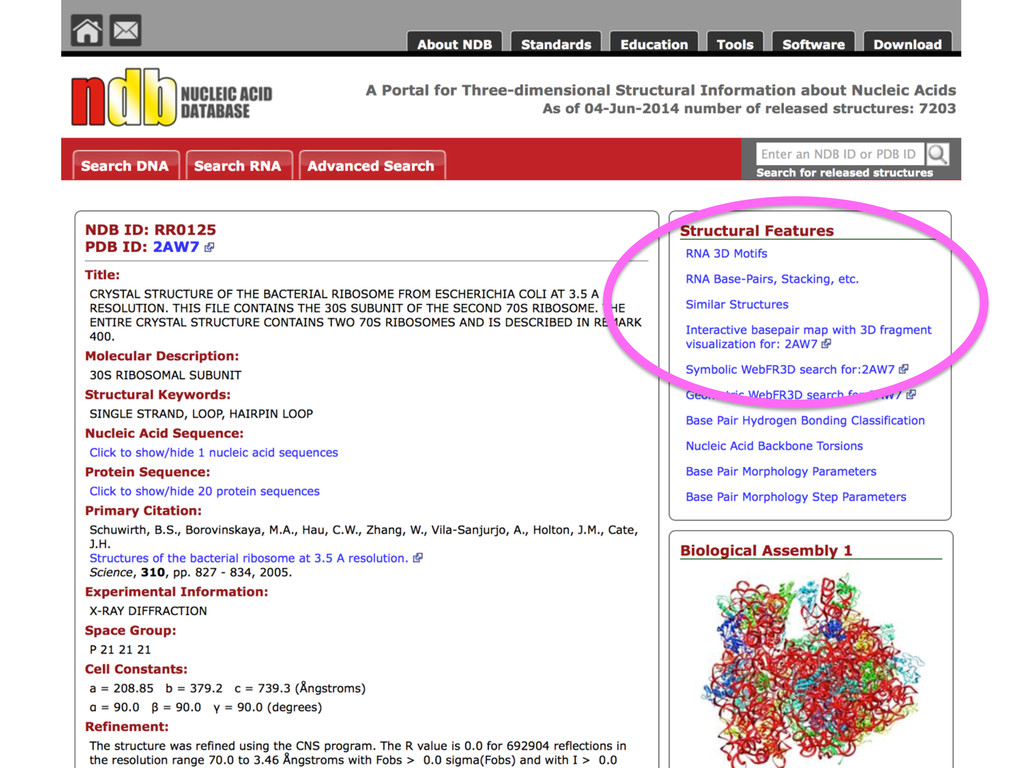
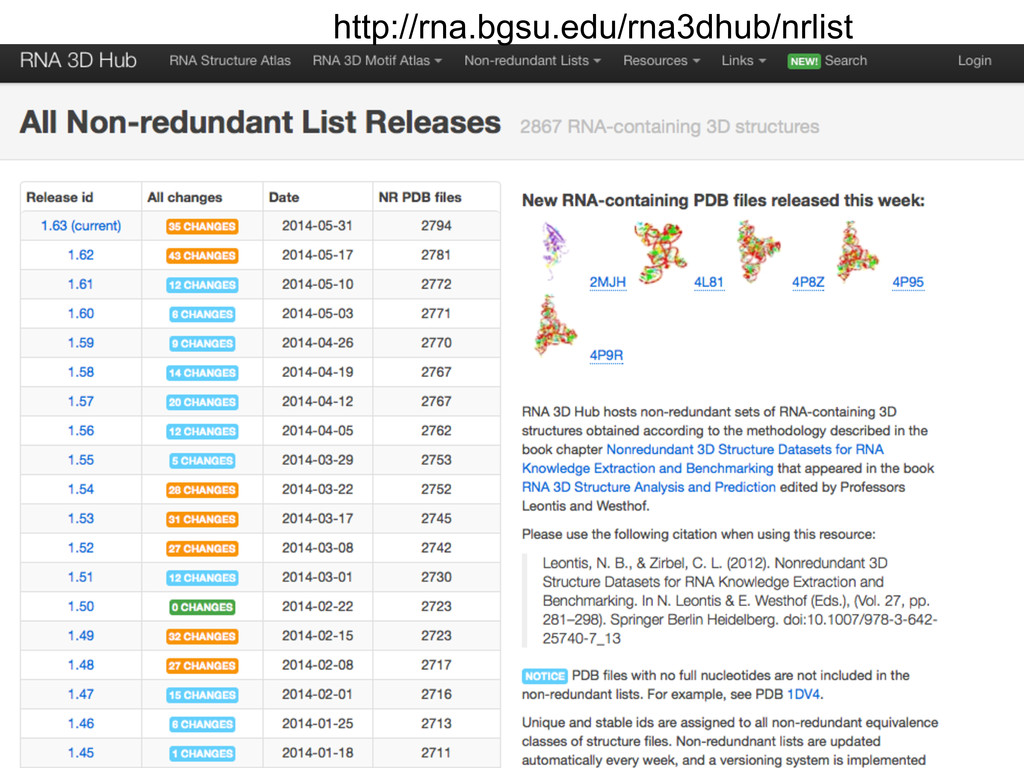
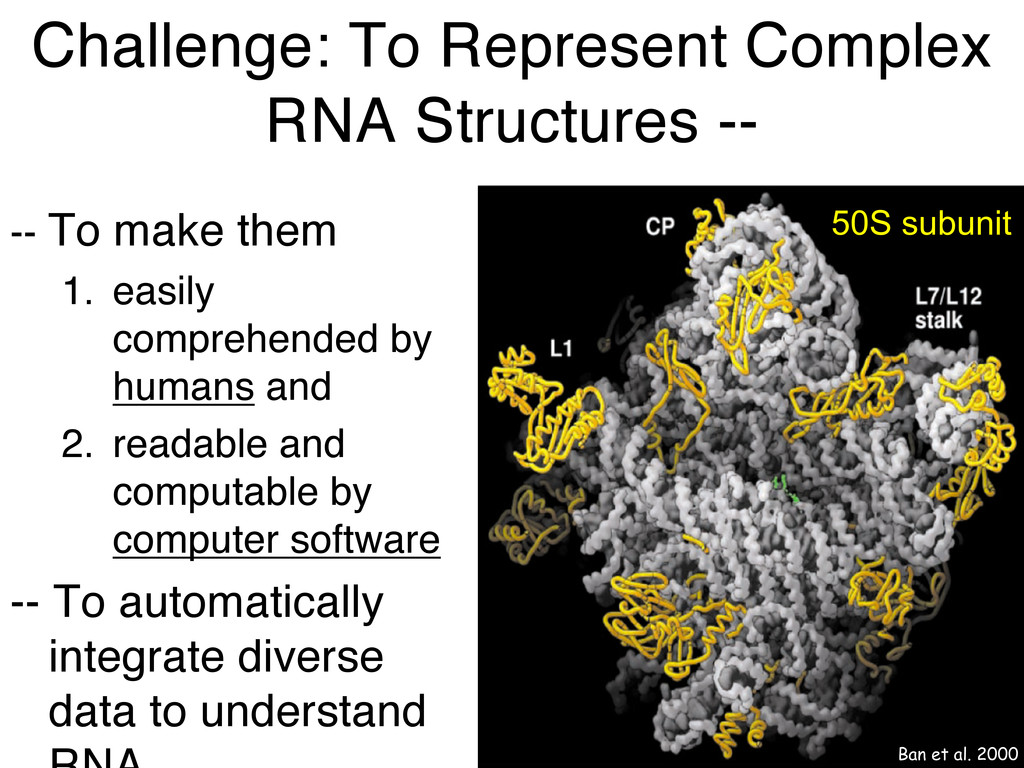
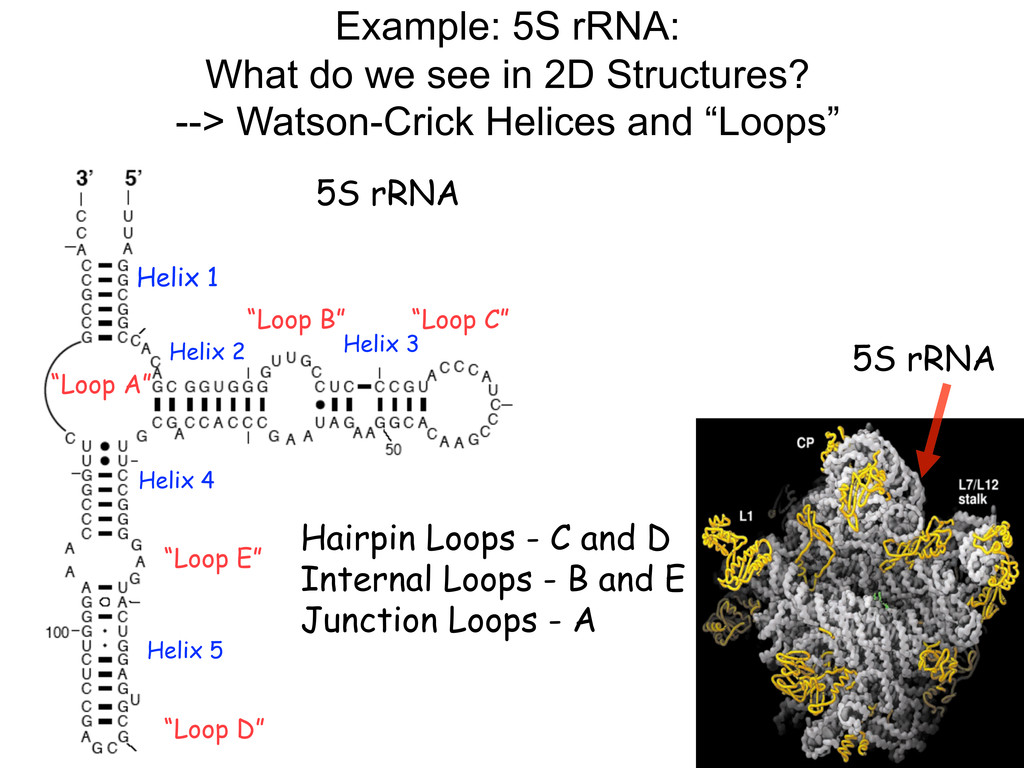
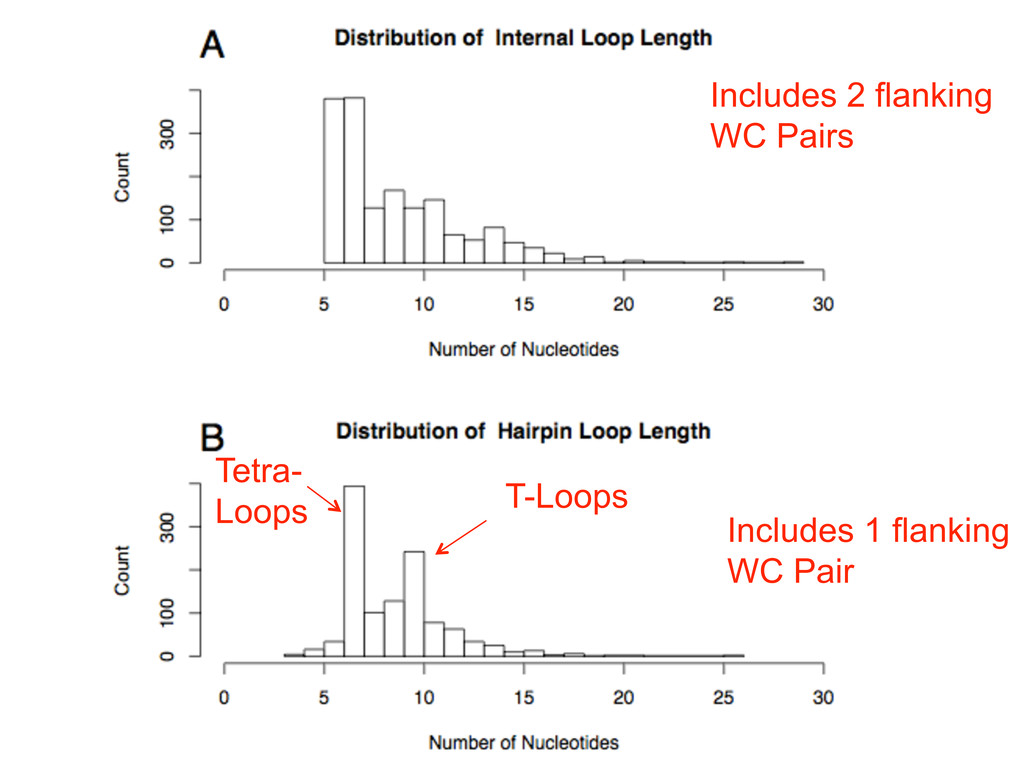
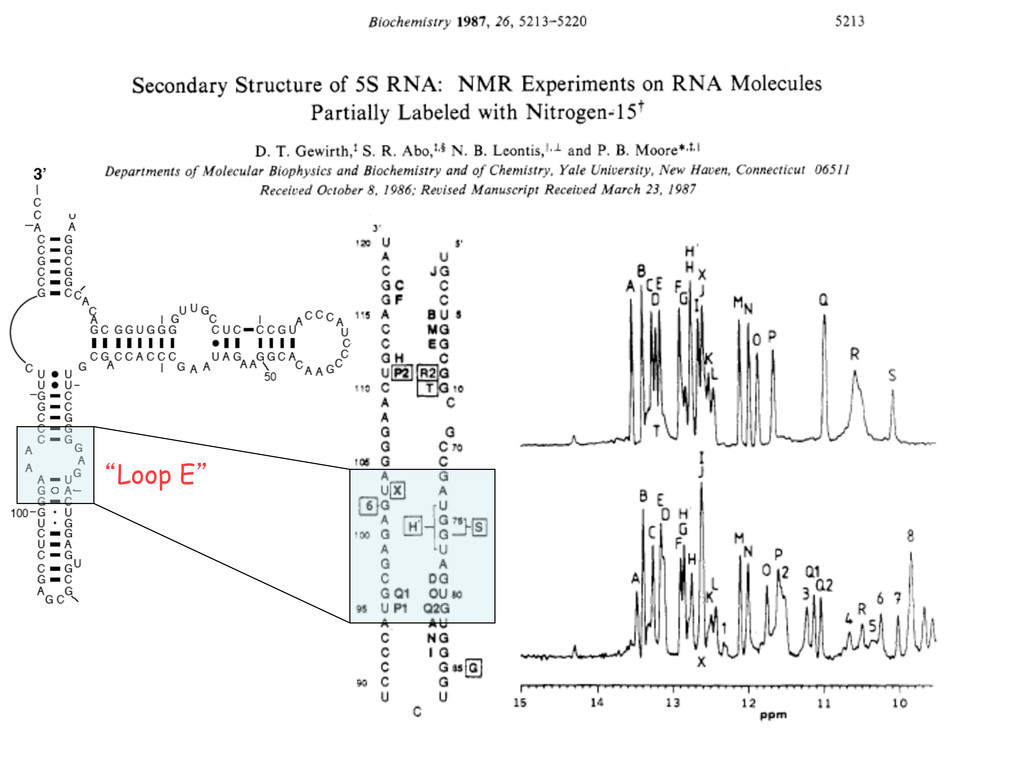
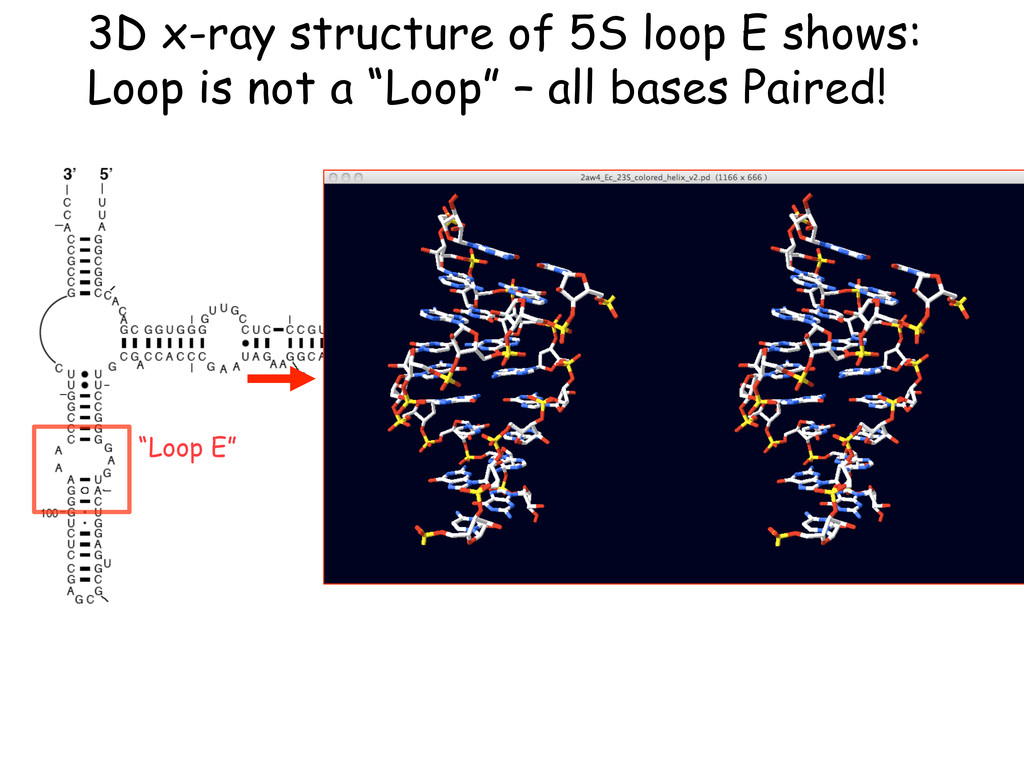
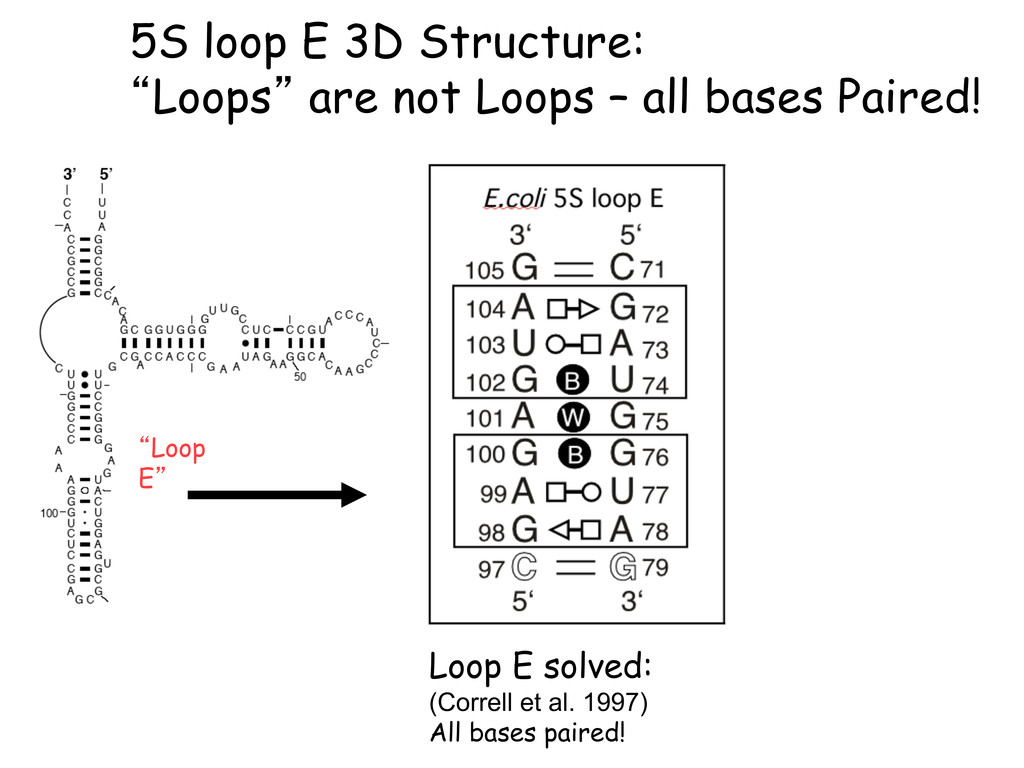
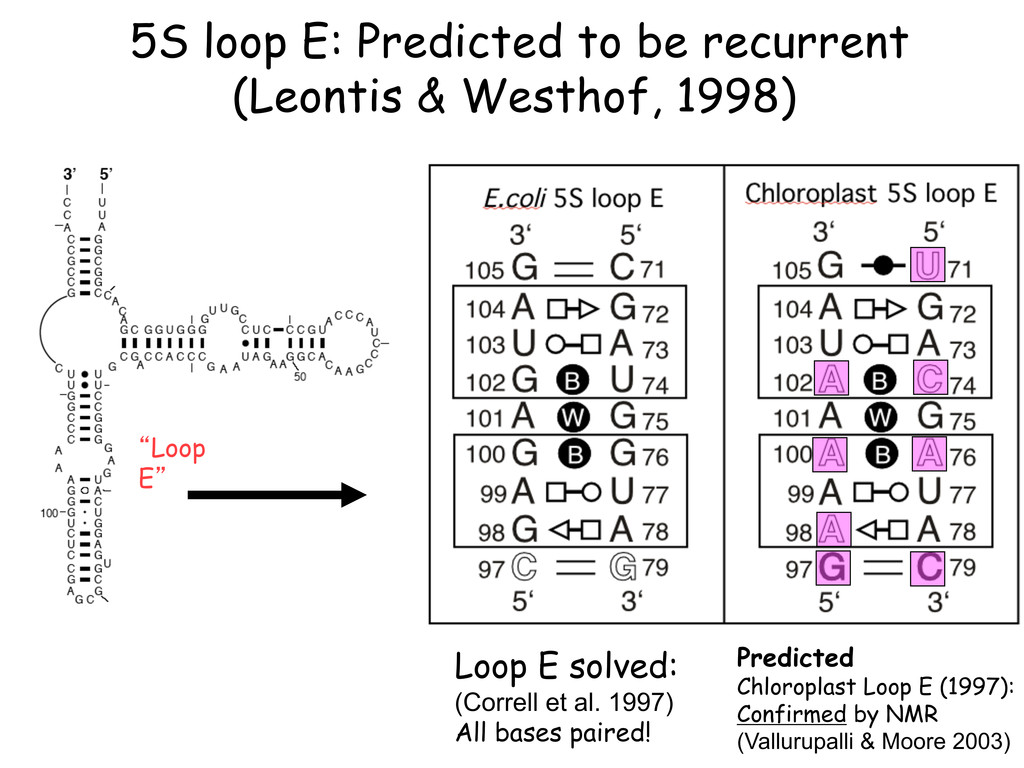
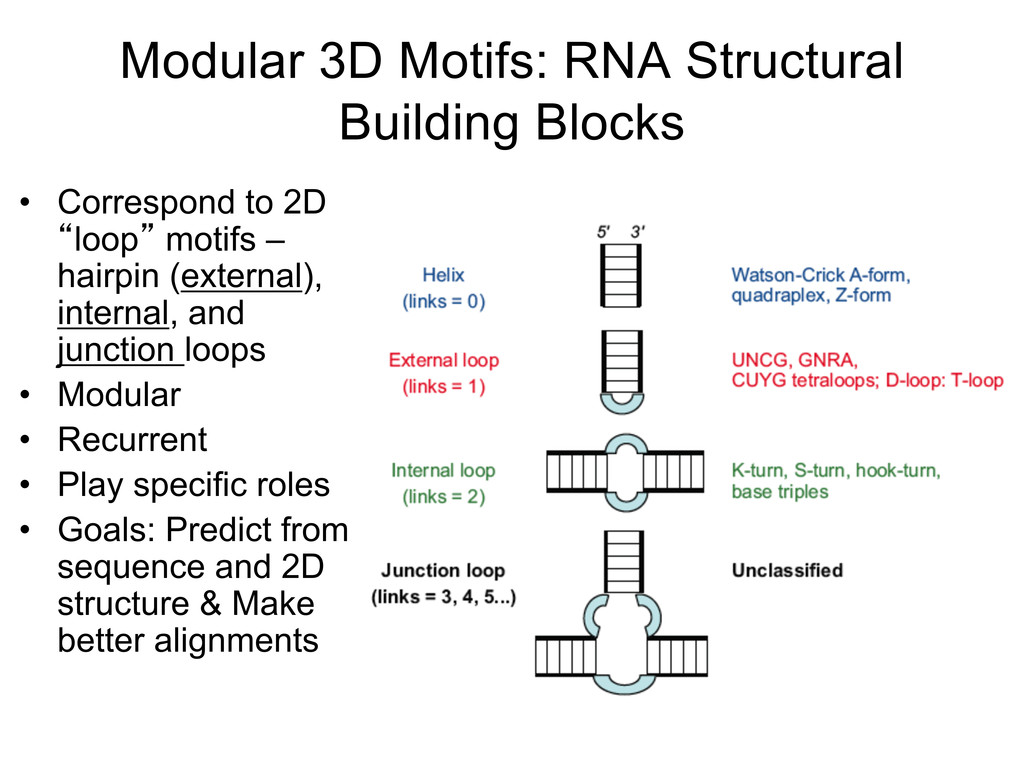
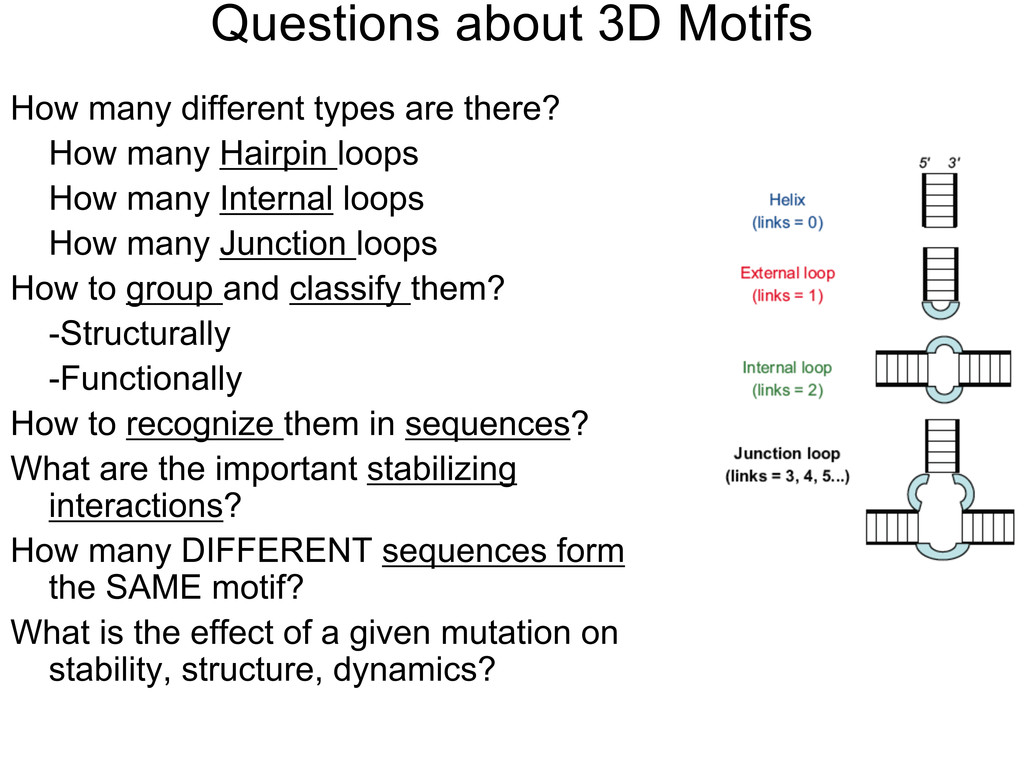
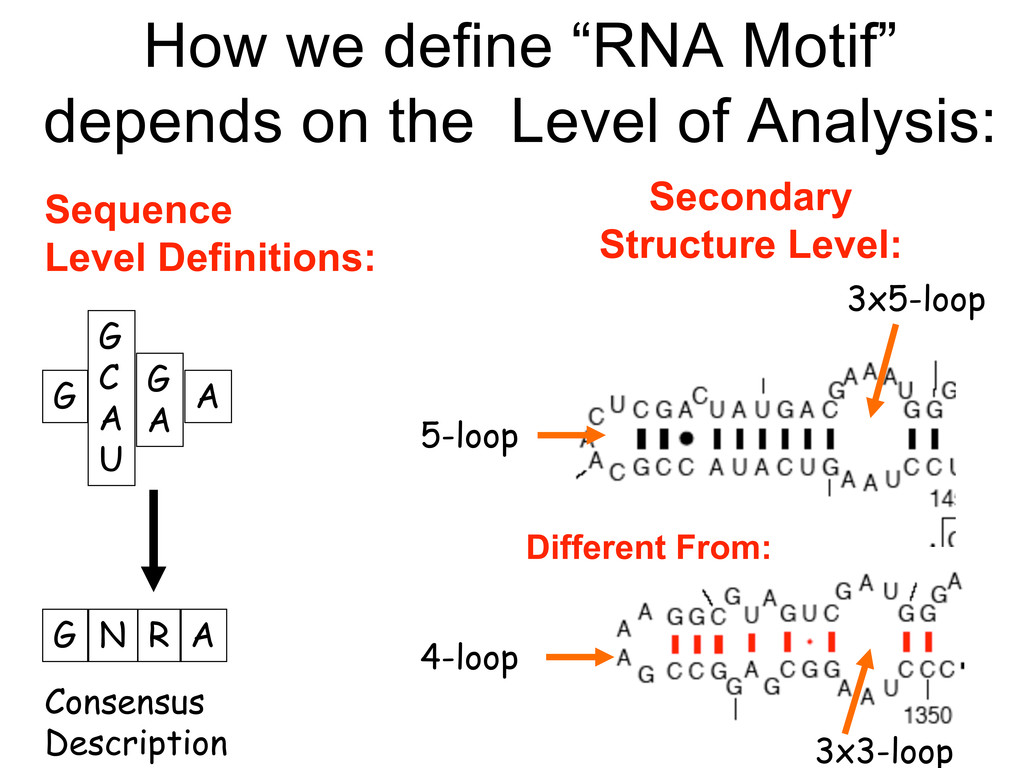
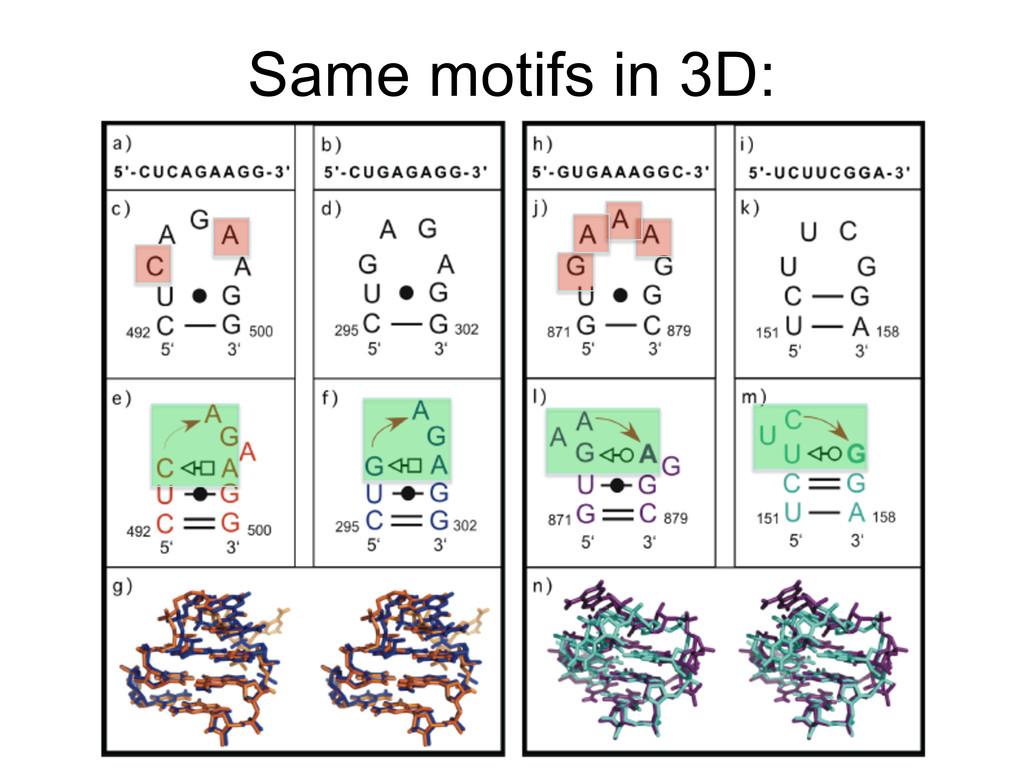
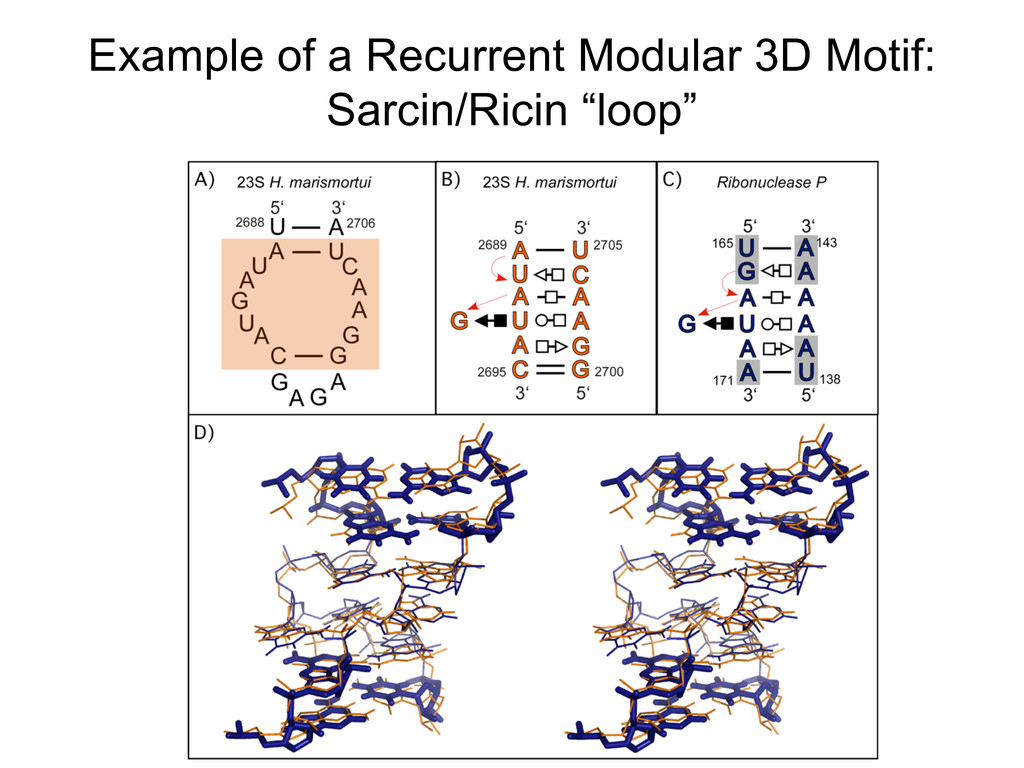
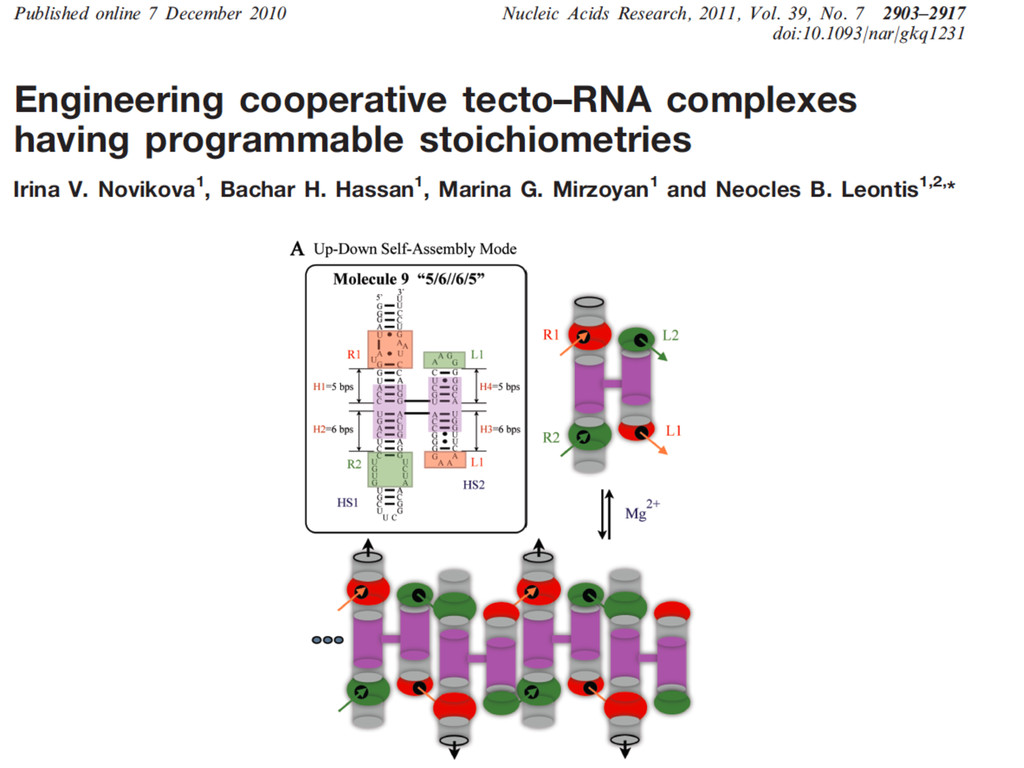
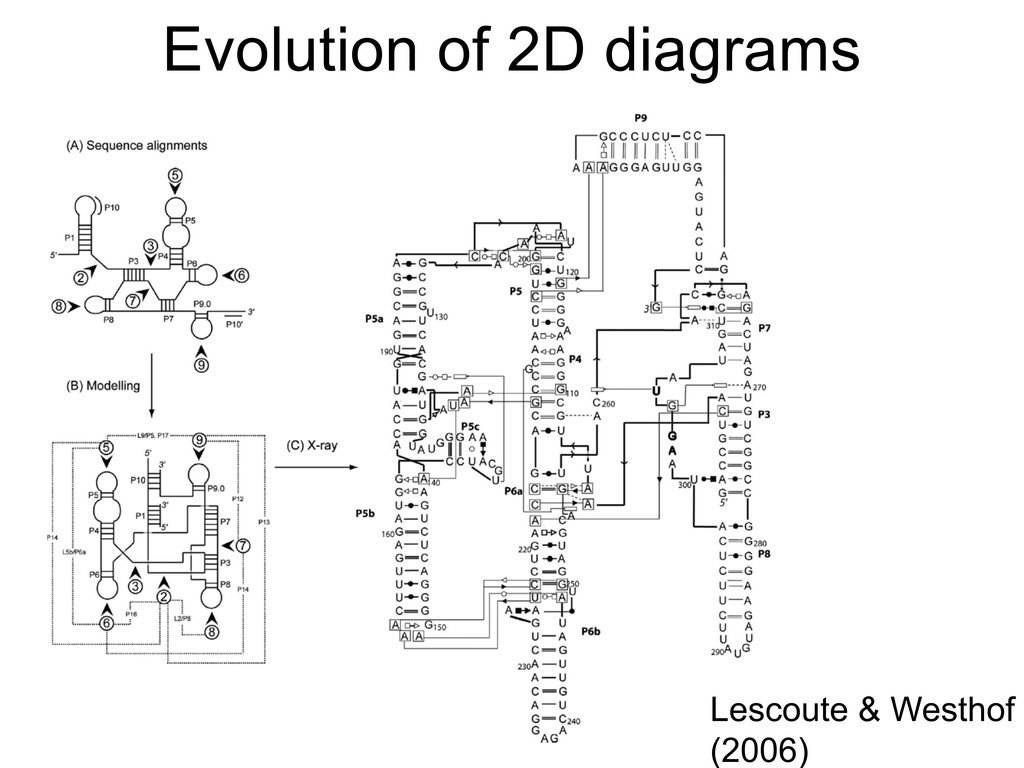
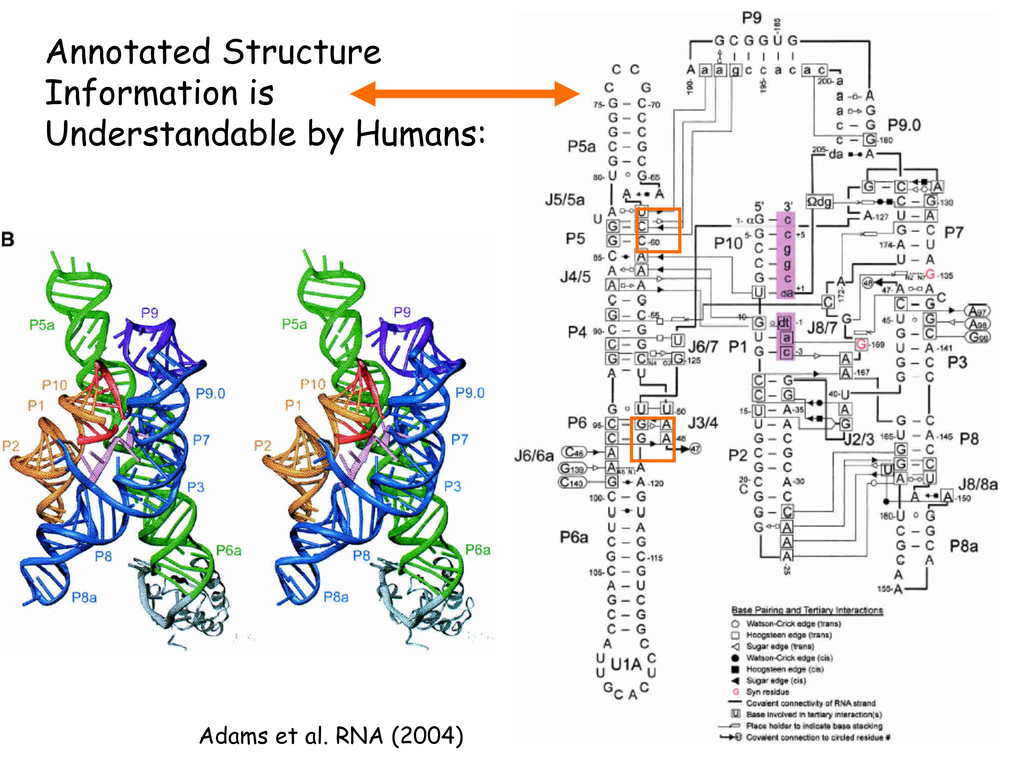
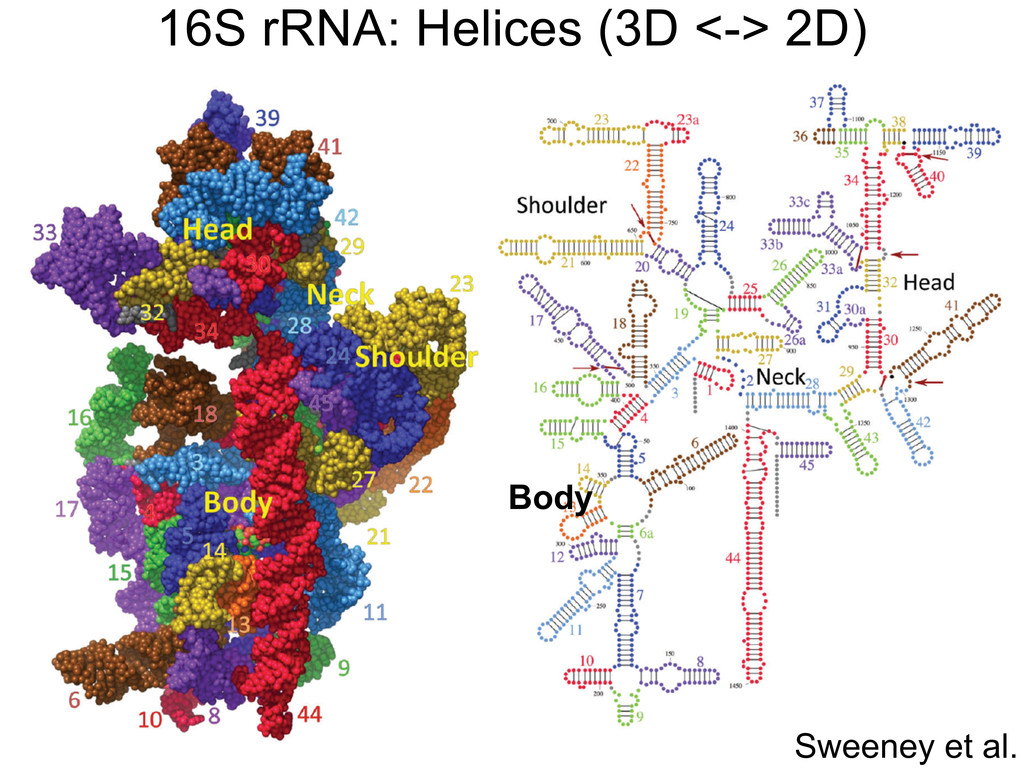
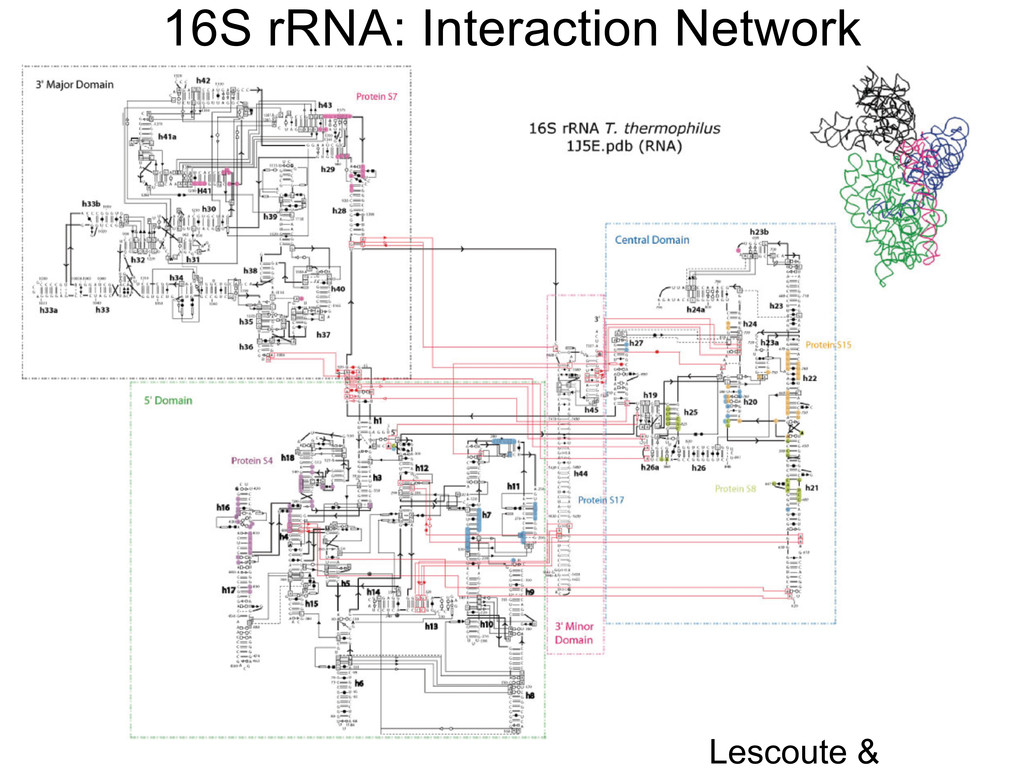
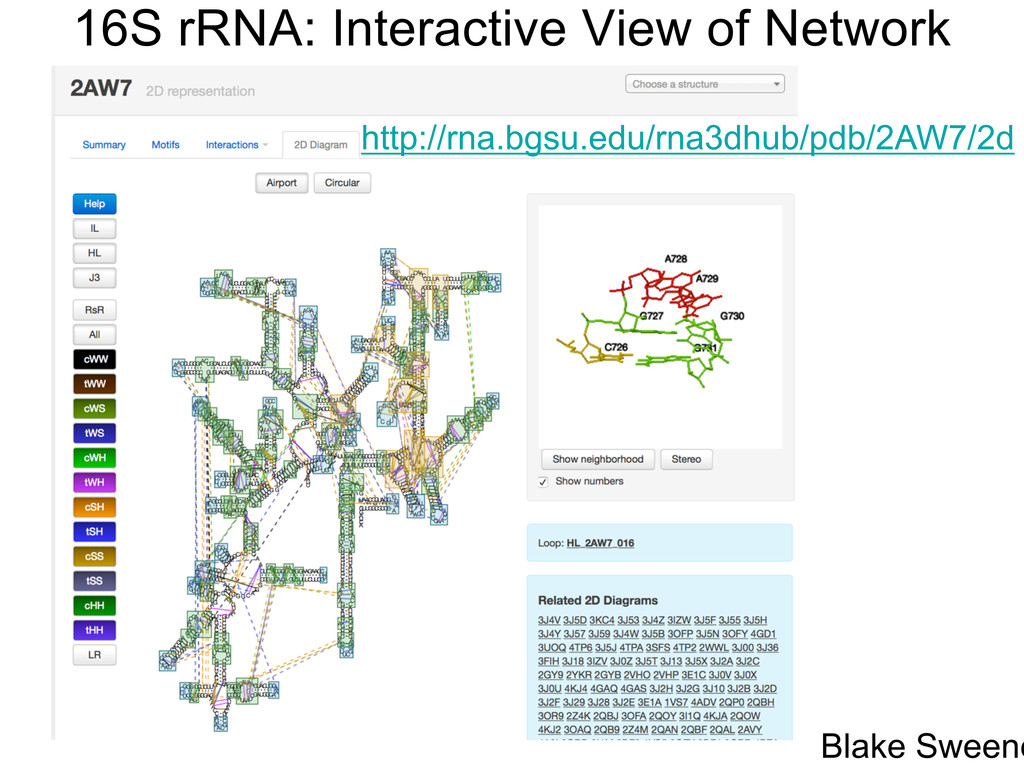
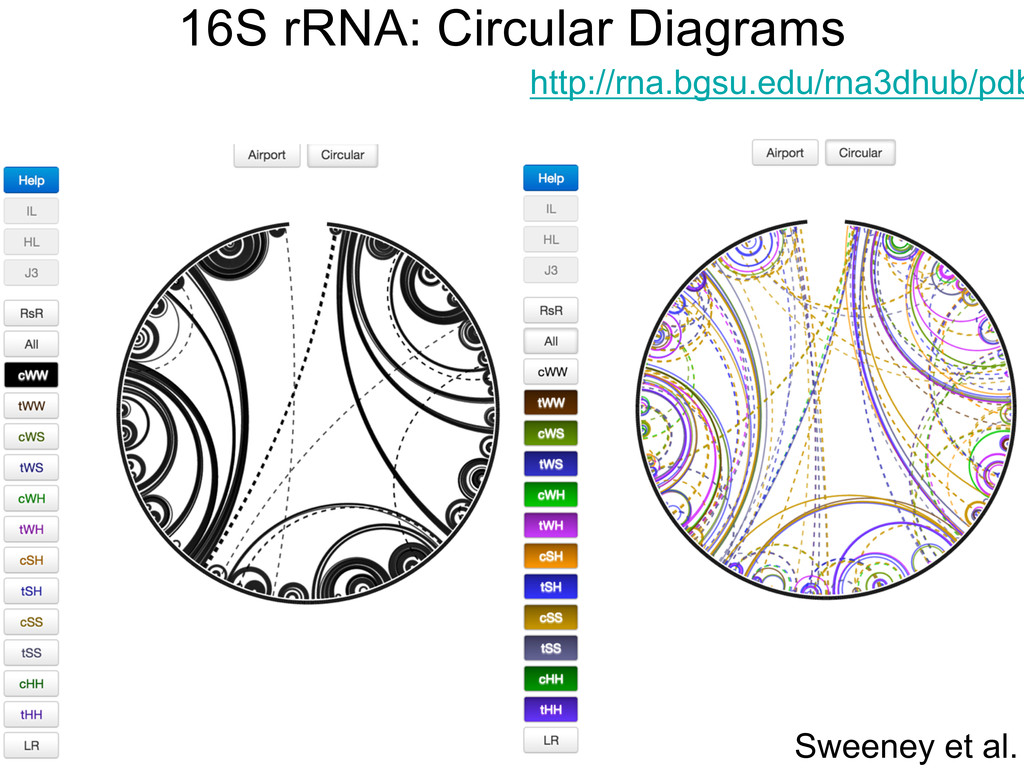
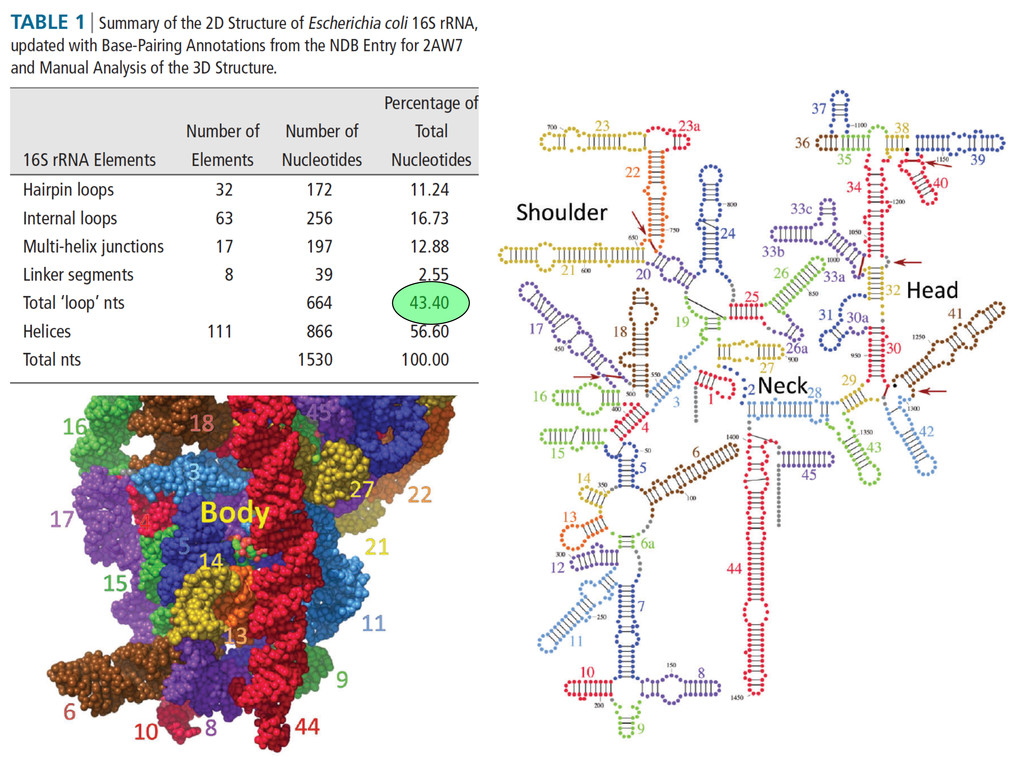
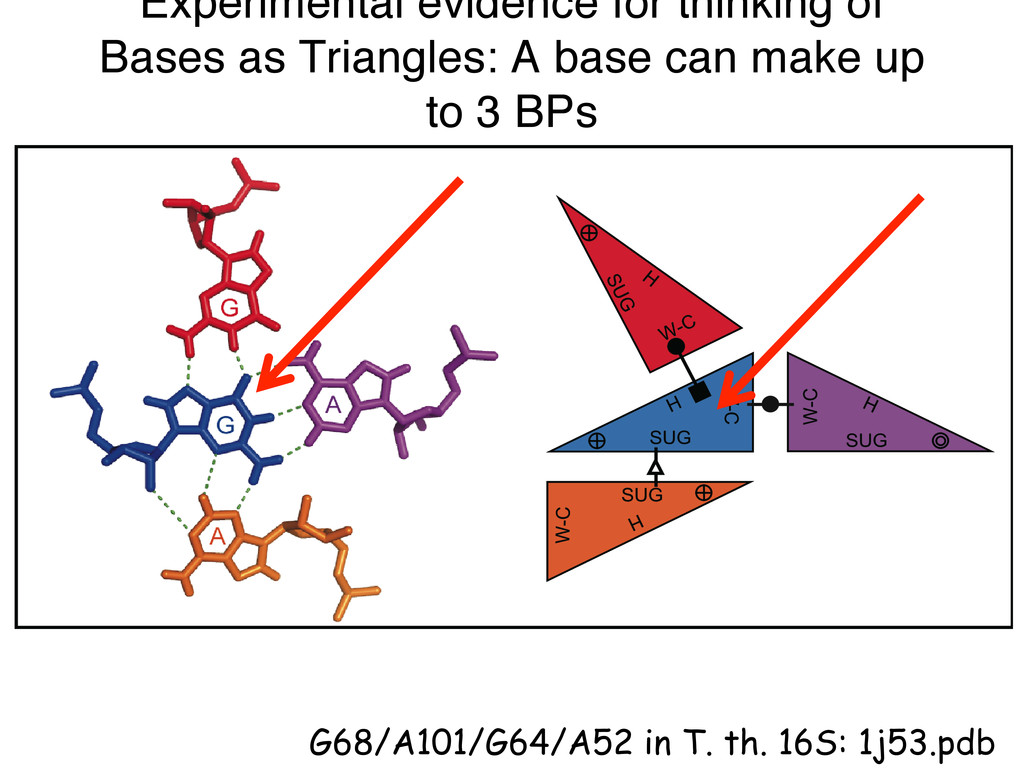
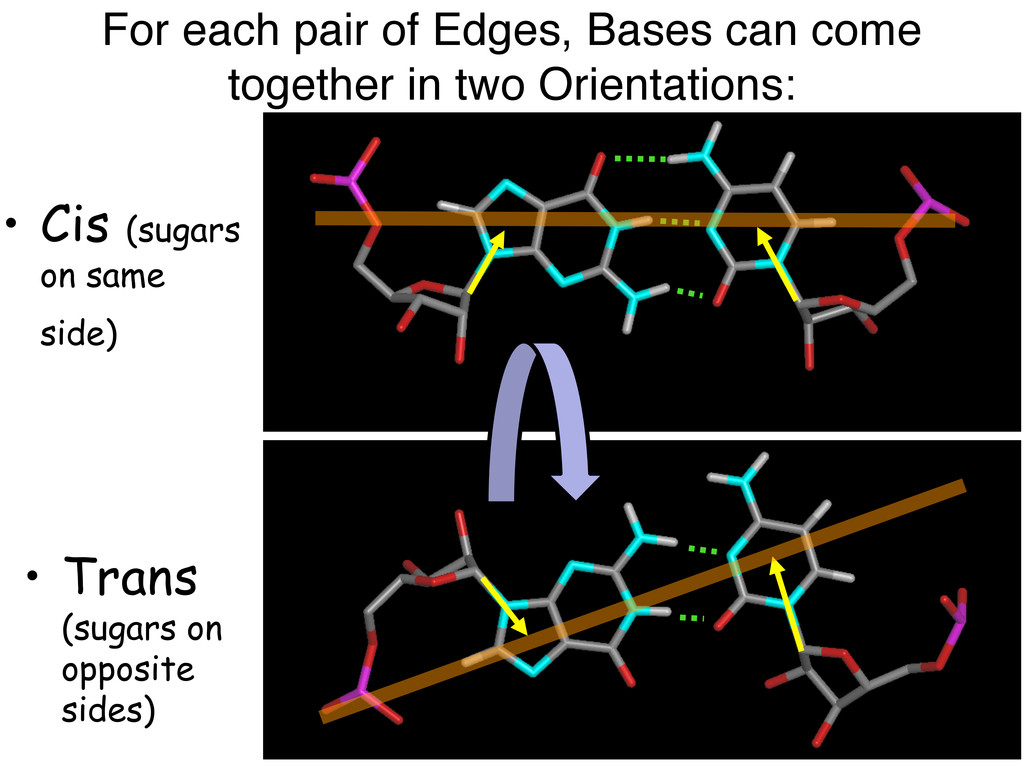
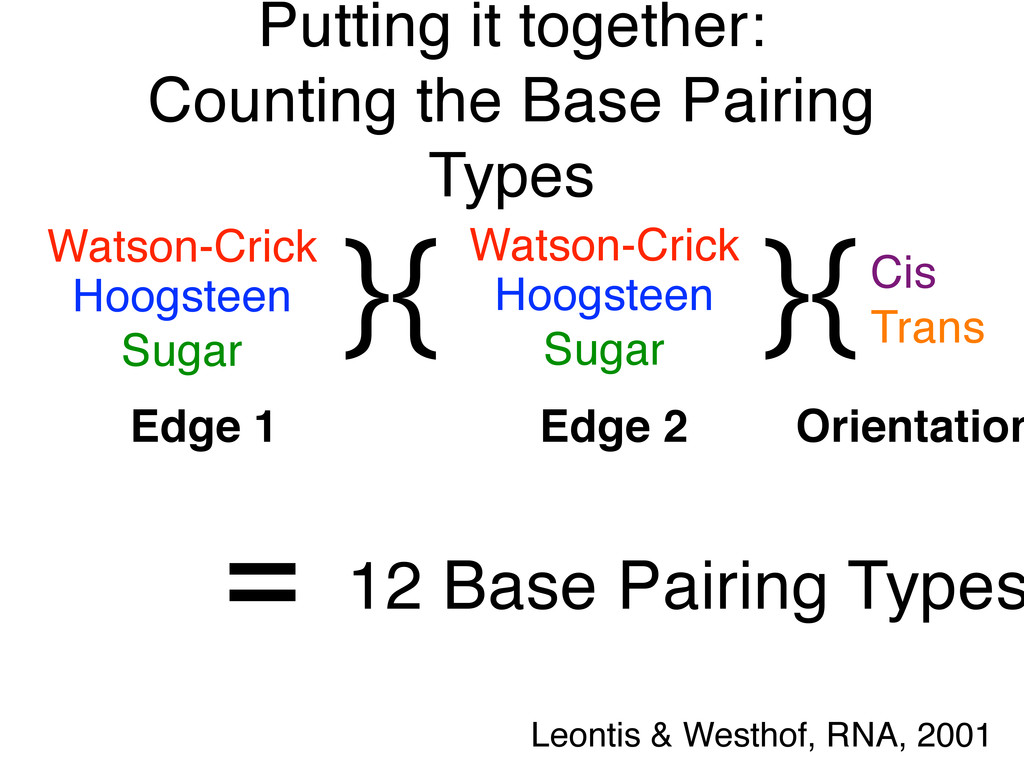
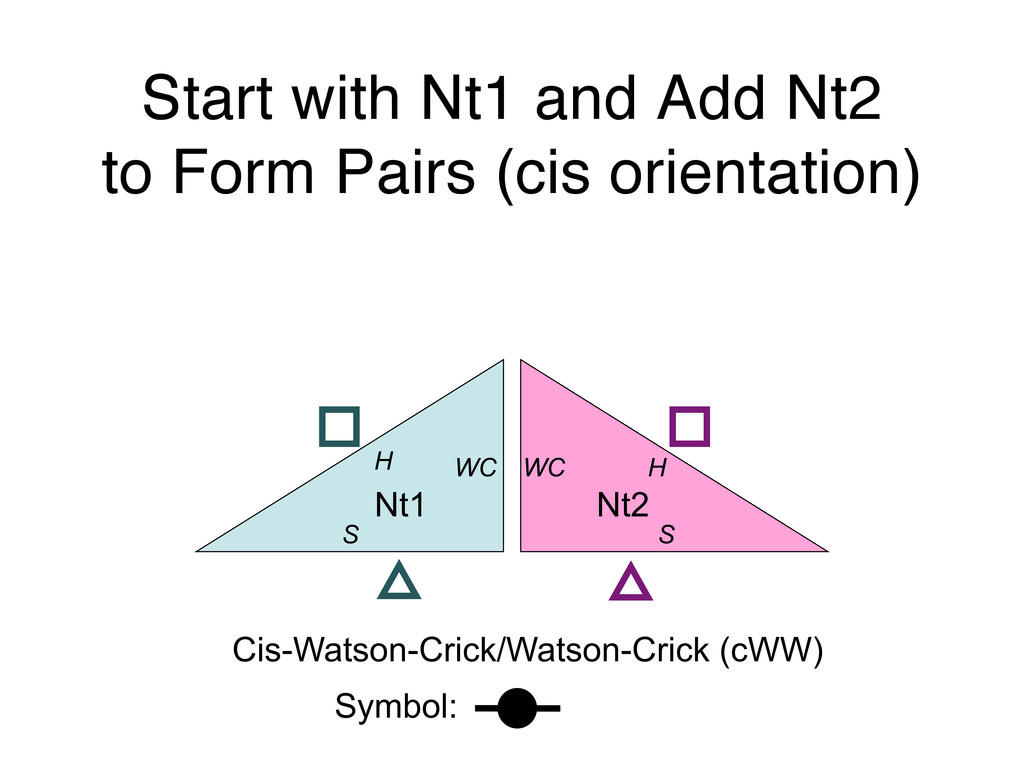
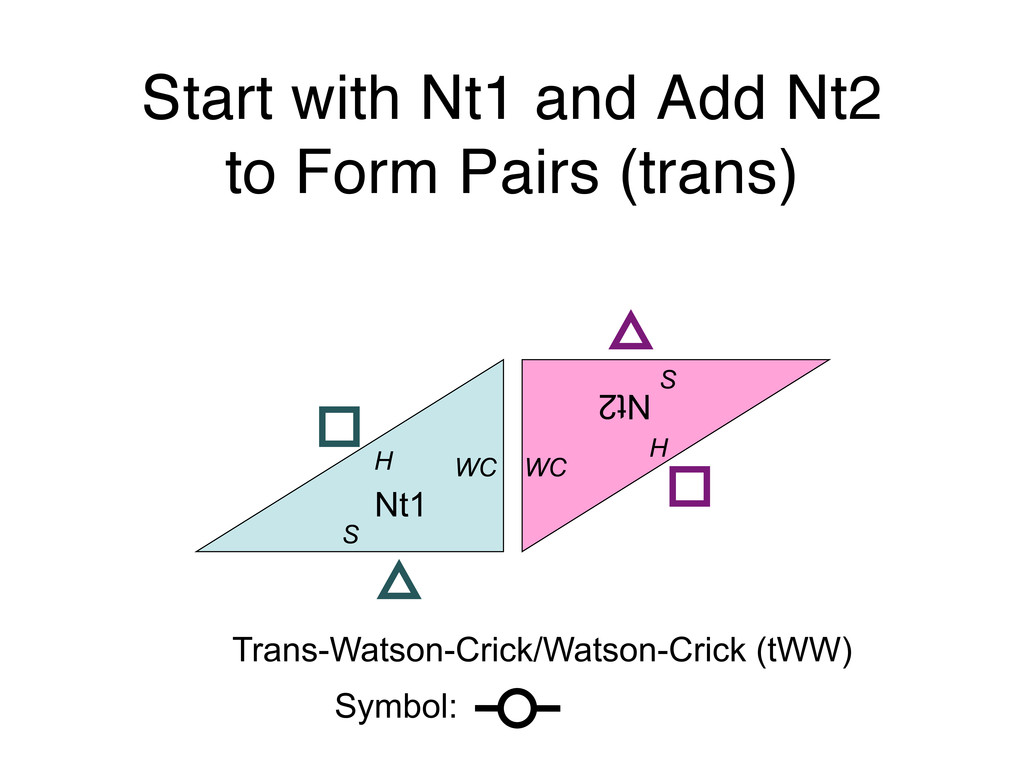
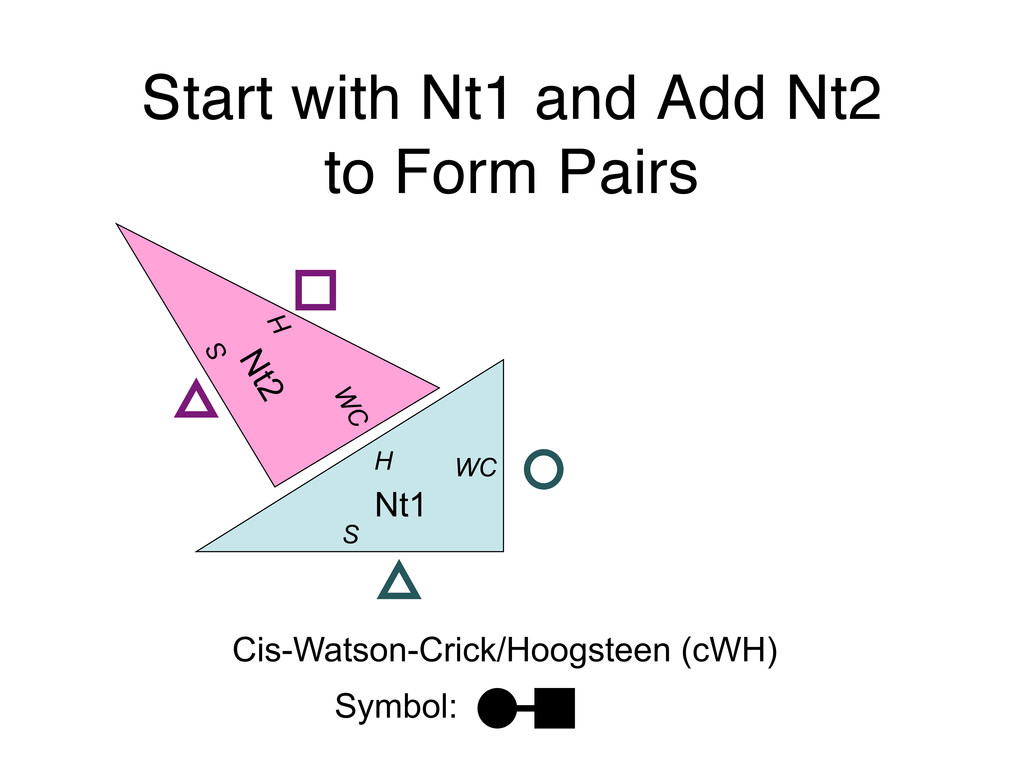
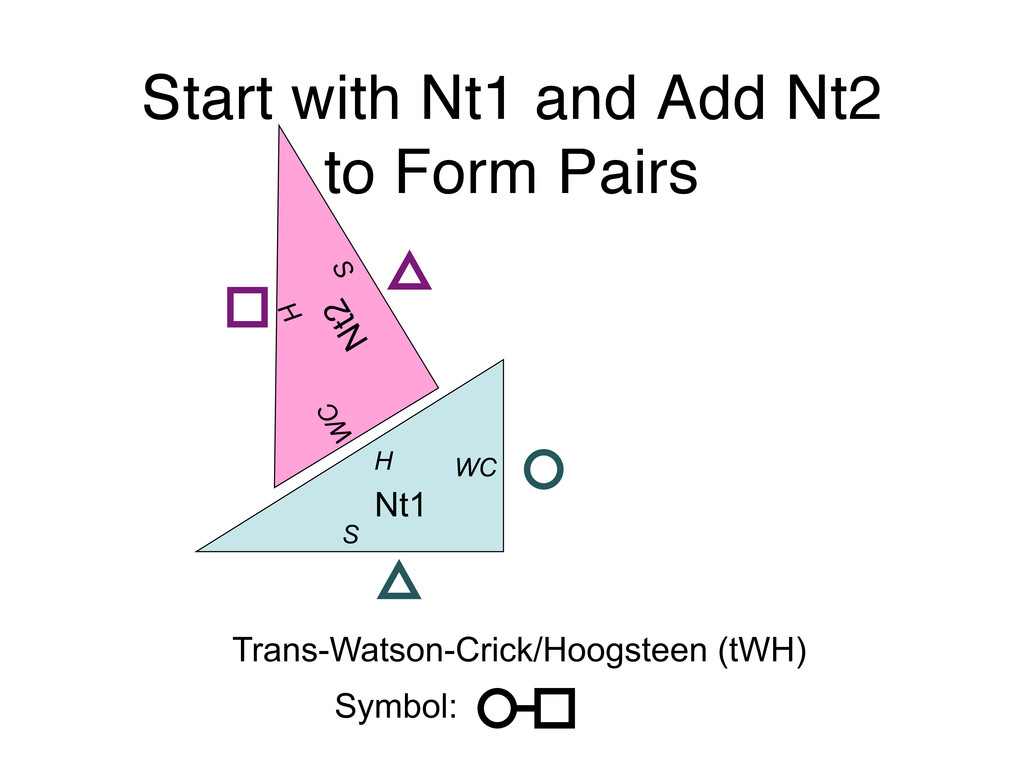
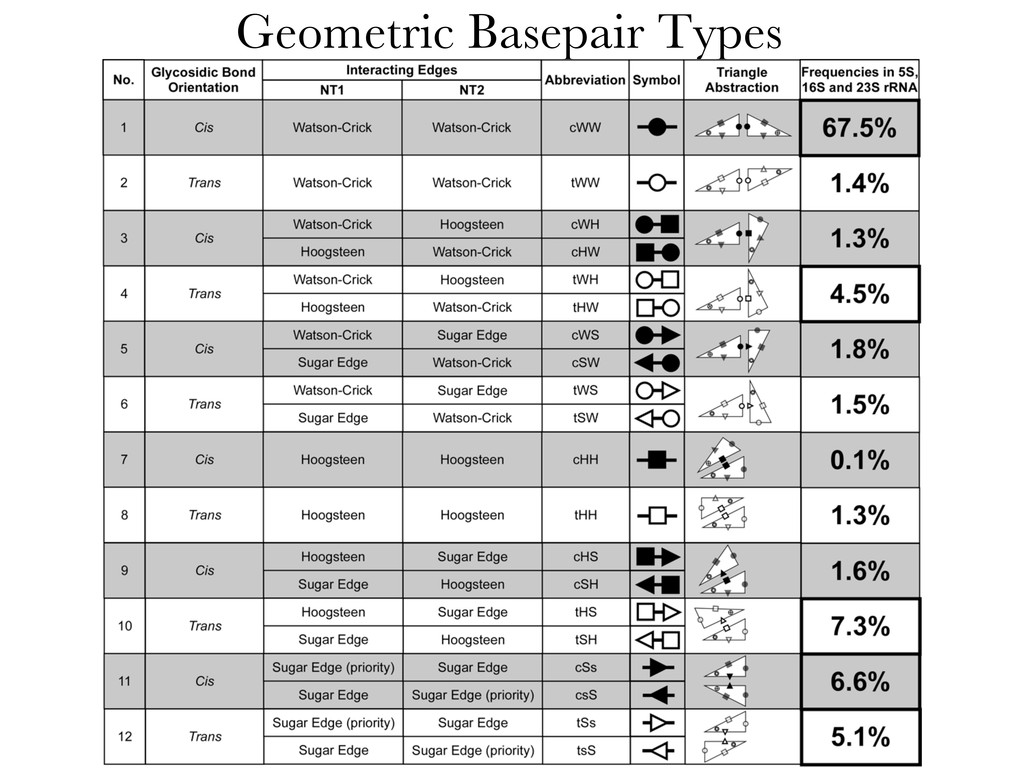
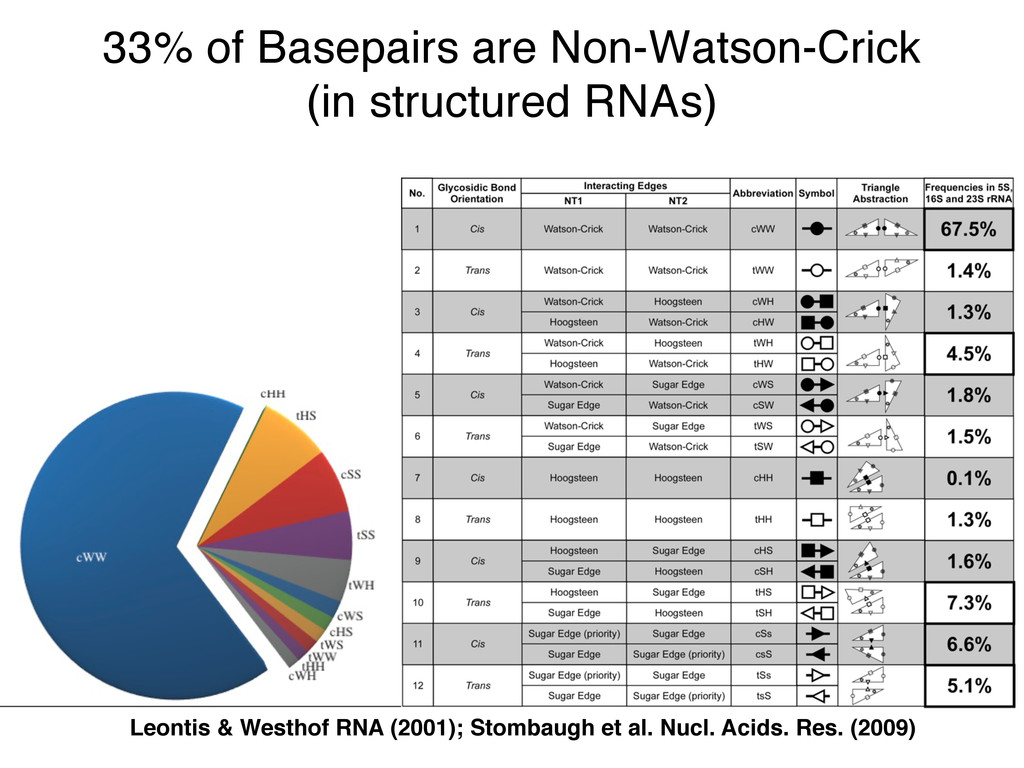
Linking and integrating diverse RNA data resources is crucial for deepening our understanding about RNA sequence, structure, function, and evolution, while also facilitating knowledge discovery for practical applications, such as RNA design for nanotechnology and nanomedicine. Atomic-resolution 3D data of highly structured RNA molecules, like ribosomal RNAs, riboswitches, and ribozymes, are rapidly accumulating. These data, appropriately integrated, are relevant for understanding many other RNA molecules since the potential for 3D structure can be as important as the sequence, even for molecules such as mRNAs, where the sequence-encoded information is central to function but structured regions can serve to modulate the translation of that information in important ways. I will use 16S rRNA as an example to illustrate the characteristics of structured RNA molecules and ways of visualizing, analyzing, and annotating RNA structures at different levels of organization, from pairwise interactions to recurrent local motifs (hairpin, internal and junction loops) to domains and folds. I will review the challenges and successes of visualising structural annotations on 2D diagrams, connecting sequence alignments with structures, and the need for automated tools to assist in comparative analysis. A major issue is integrated access to the different forms of RNA data by diverse users in ways that facilitate knowledge discovery. A key challenge is that their use is currently limited by the familiarity of users with the specialized software tools for accessing, interpreting, and assessing the quality of those data. Ways are needed to overcome these barriers to provide actionable information to diverse users regardless of their scientific backgrounds.
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
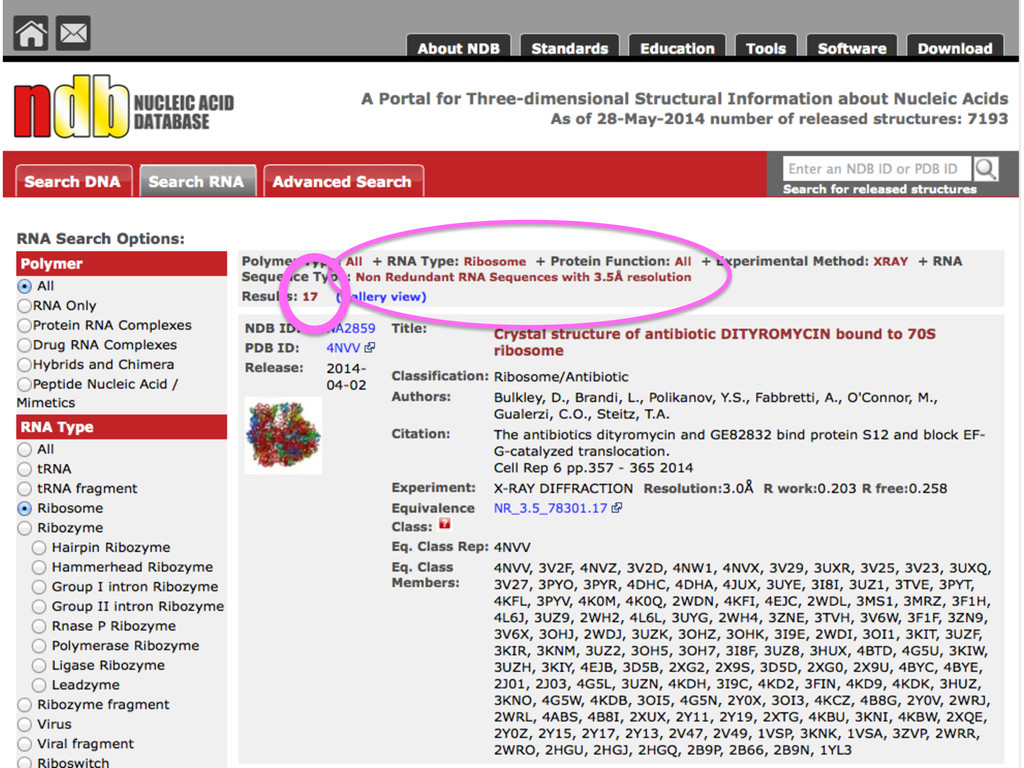{kind=link}
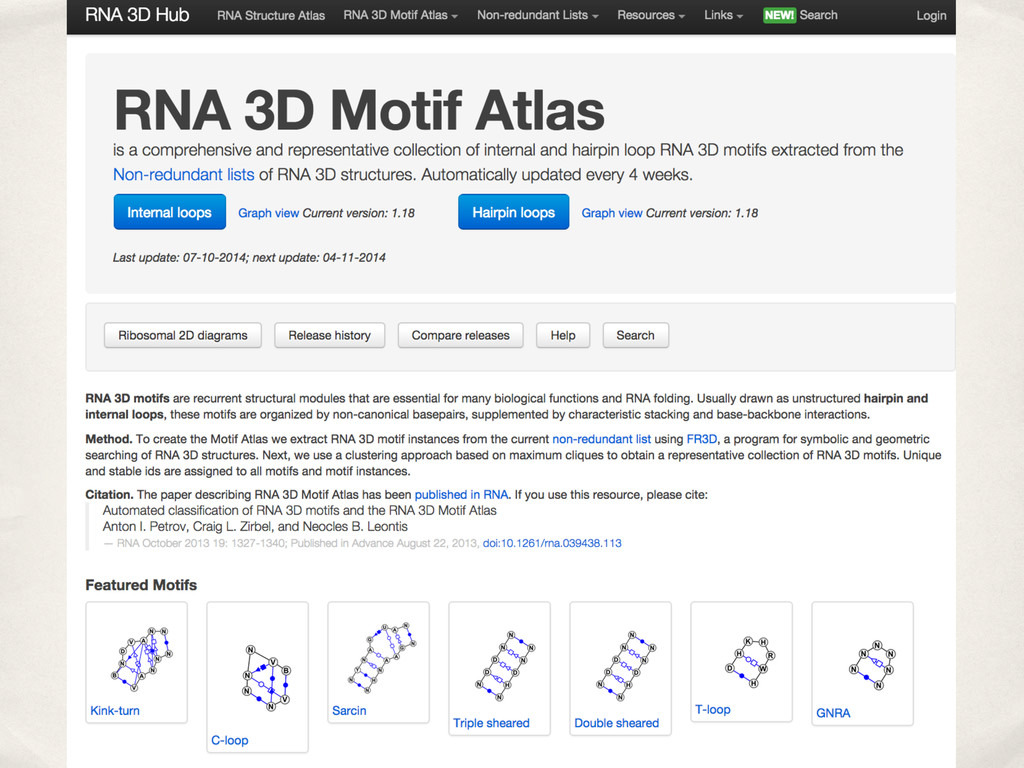{kind=link}
{kind=link}
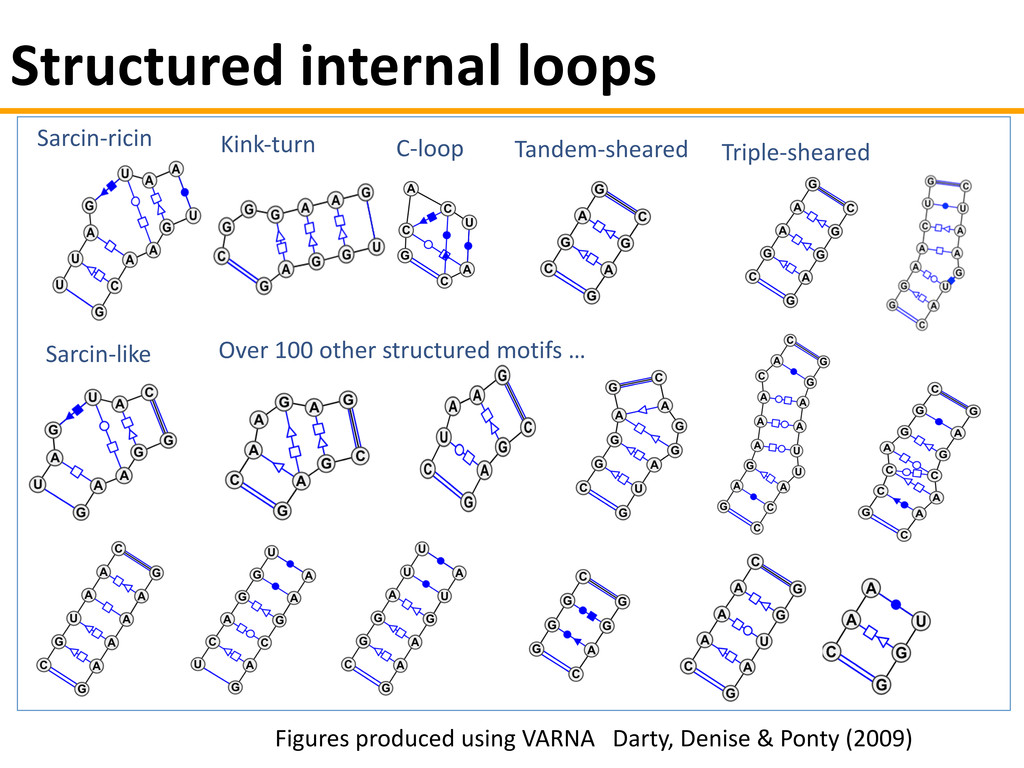{kind=link}
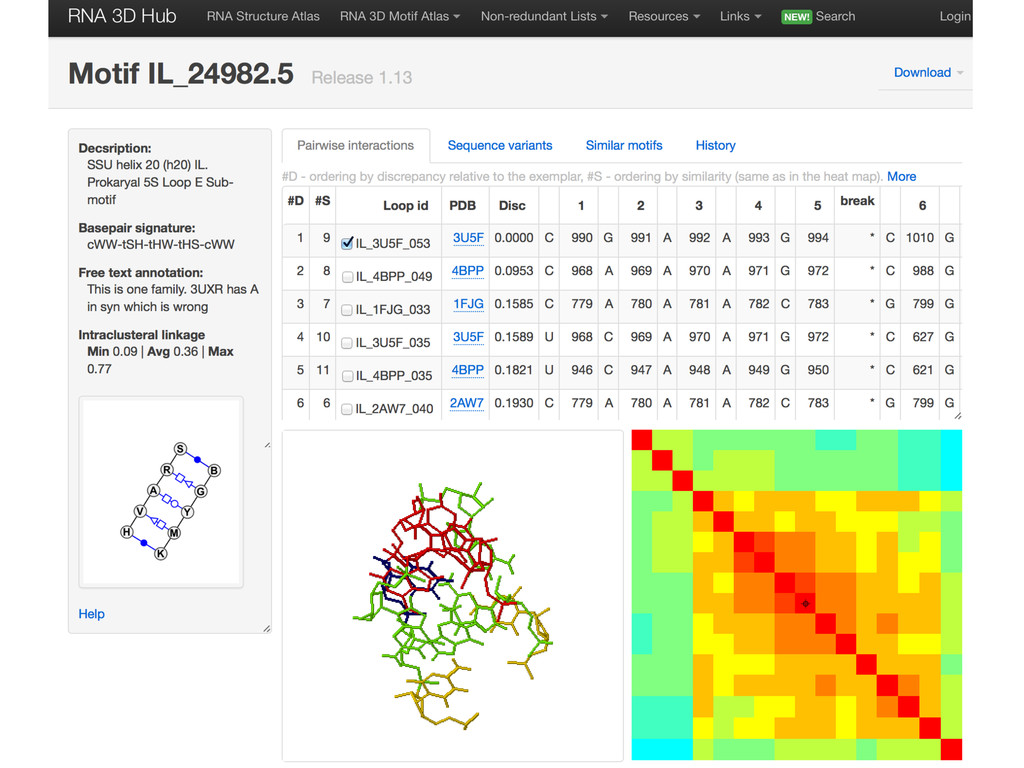{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
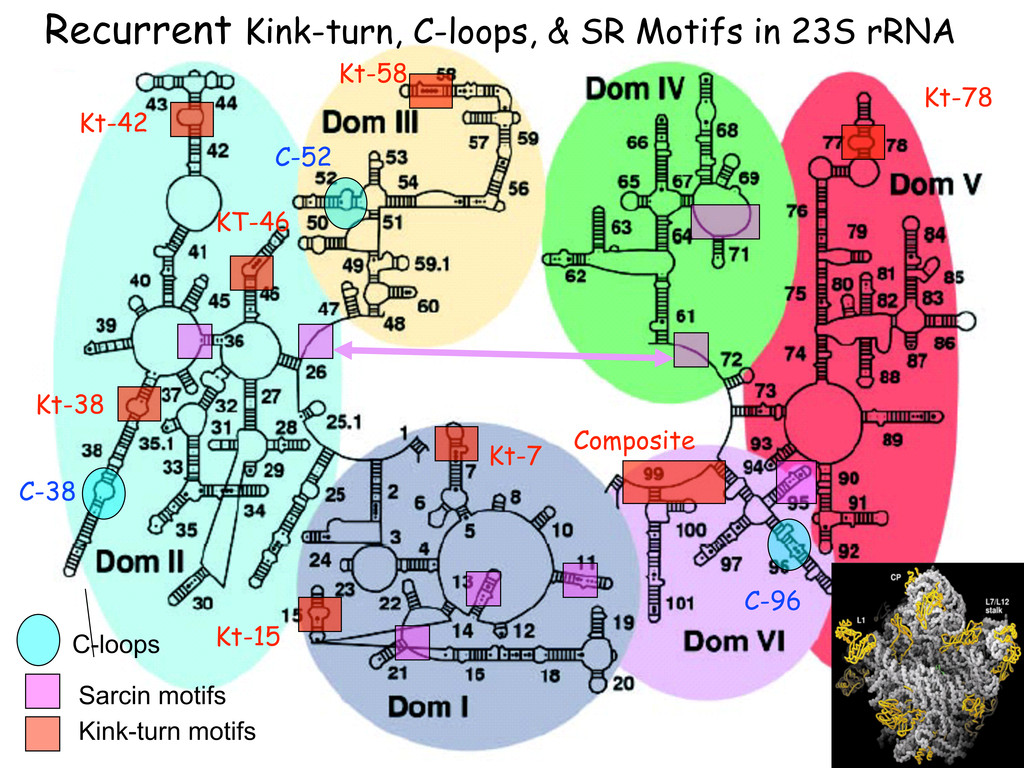{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
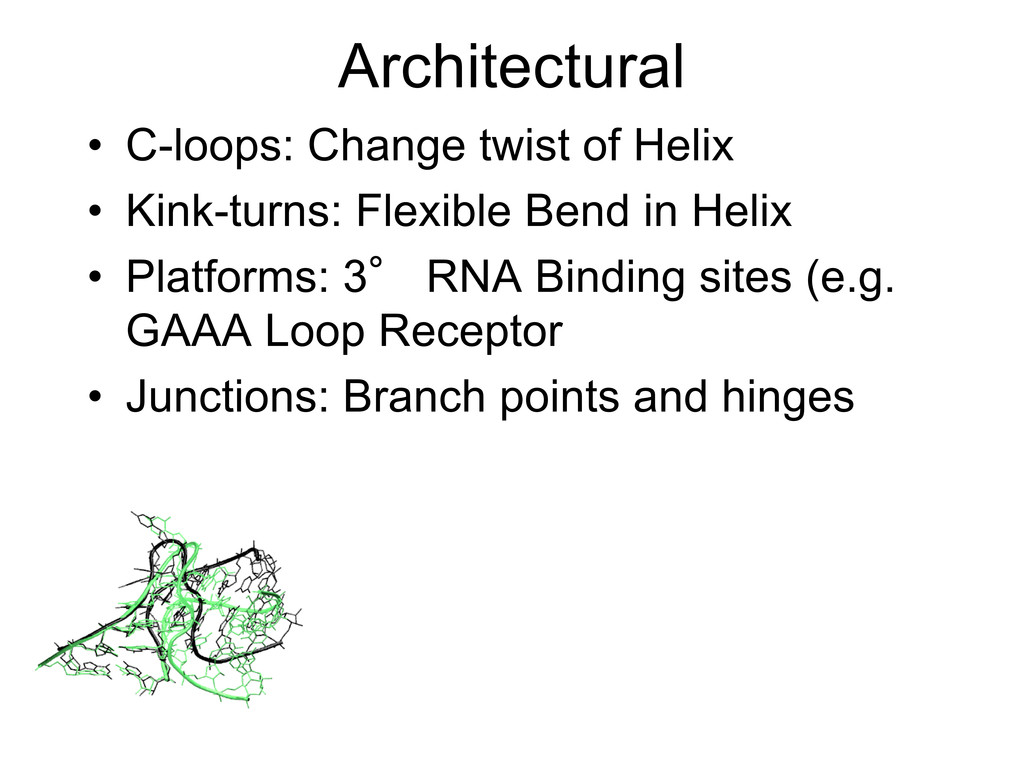{kind=link}
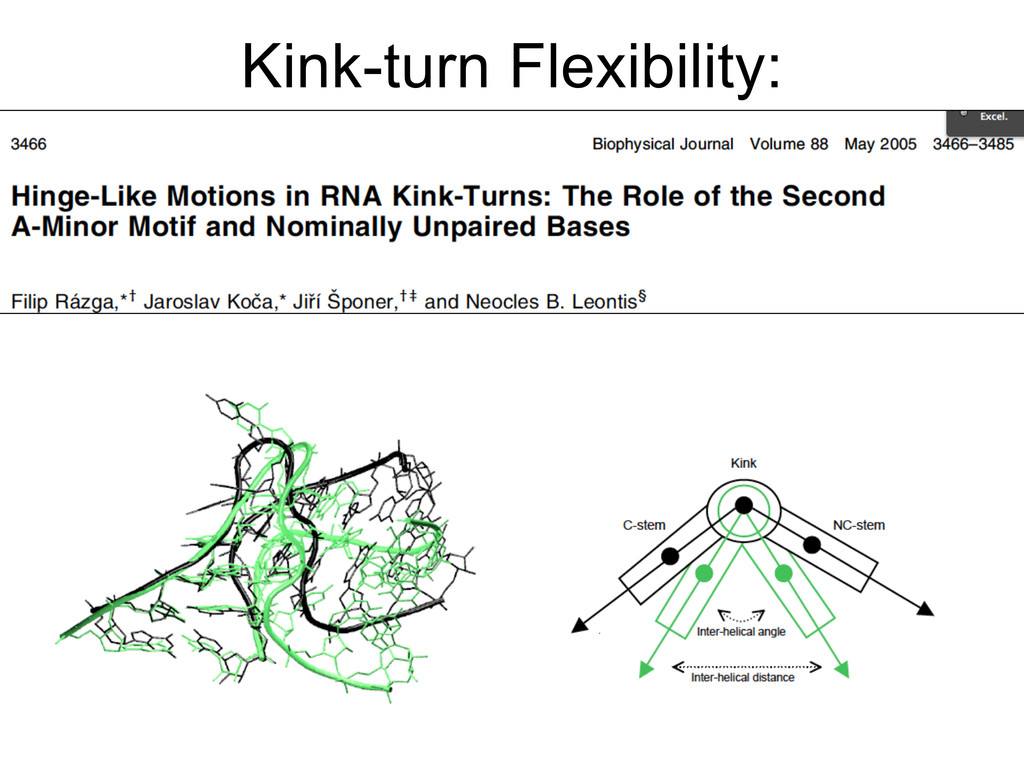{kind=link}
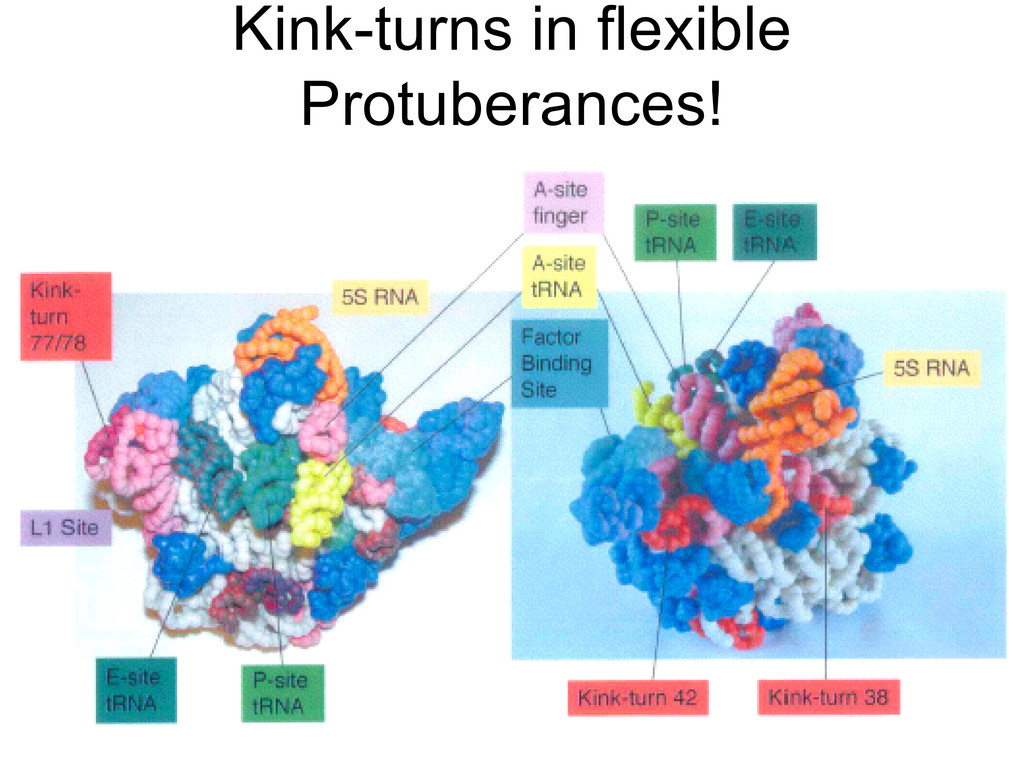{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}