Slides summarizing the first chapter of Malosree Maitra's Thesis "Cell type and sex specific transcriptomics of the dorsolateral prefrontal cortex in depression" presented in the libd_lcolladotor_team weekly seminar (02_14_2024).
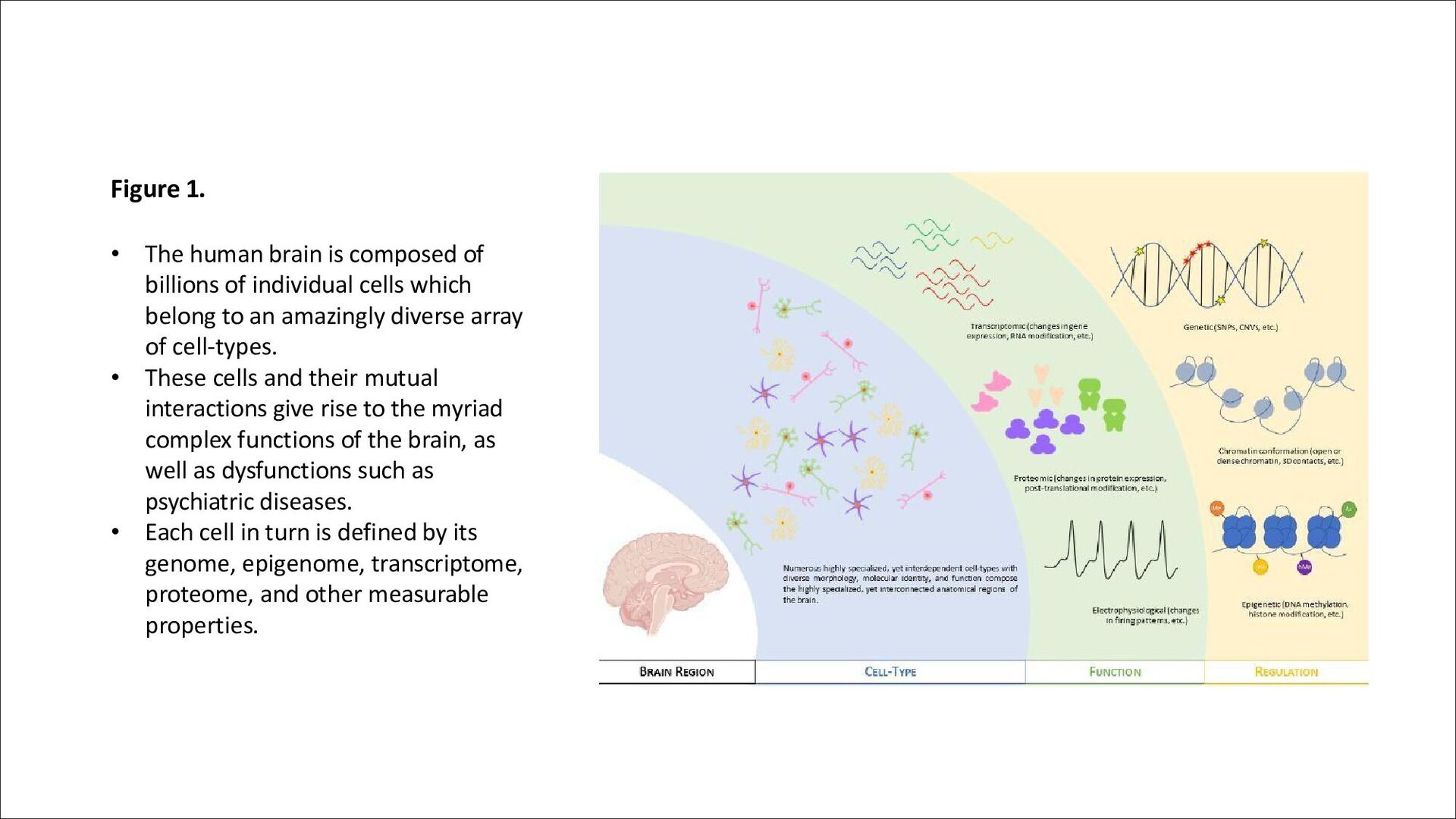
of individual cells which belong to an amazingly diverse array of cell-types. • These cells and their mutual interactions give rise to the myriad complex functions of the brain, as well as dysfunctions such as psychiatric diseases. • Each cell in turn is defined by its genome, epigenome, transcriptome, proteome, and other measurable properties.
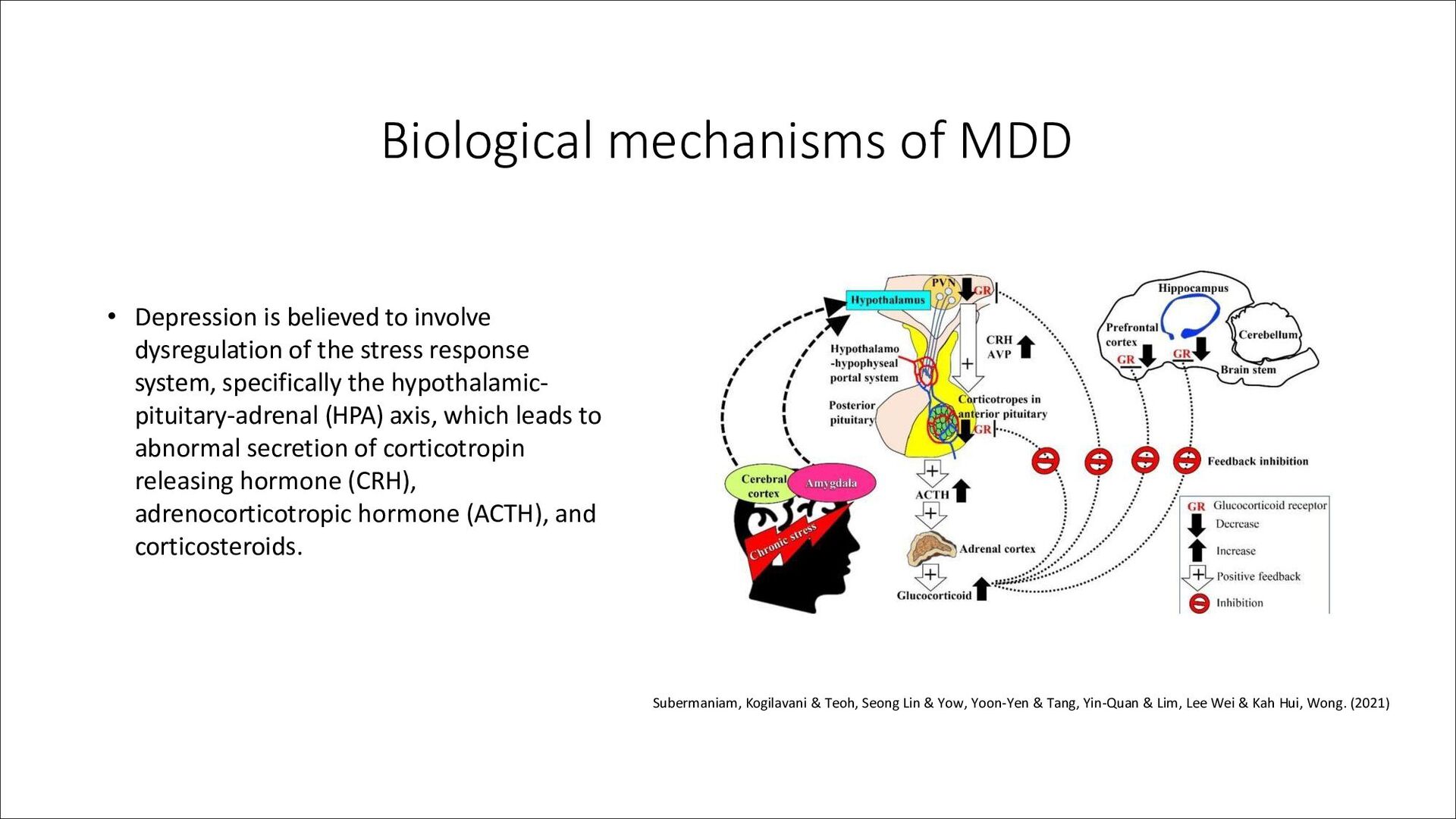
characterized by persistent low mood, anhedonia, feelings of guilt and worthlessness, and suicidal ideation among other symptoms. • It affects over 200 million people globally and is a leading cause of disability worldwide. • Depression is heterogeneous in its presentation, with varying levels of symptom severity, recurrence, treatment resistance, and associated comorbidities.
heritability (around 30-40%). Recent GWAS have identified over 100 genetic loci associated with the disorder. • Brain imaging studies indicate region-specific changes in brain activity and morphology in depression, and brain region-specific transcriptomic and epigenetic changes have been reported in post-mortem studies. • The incidence of depression is higher in women than in men, and there is accumulating evidence that the molecular basis of depression may differ between the sexes.
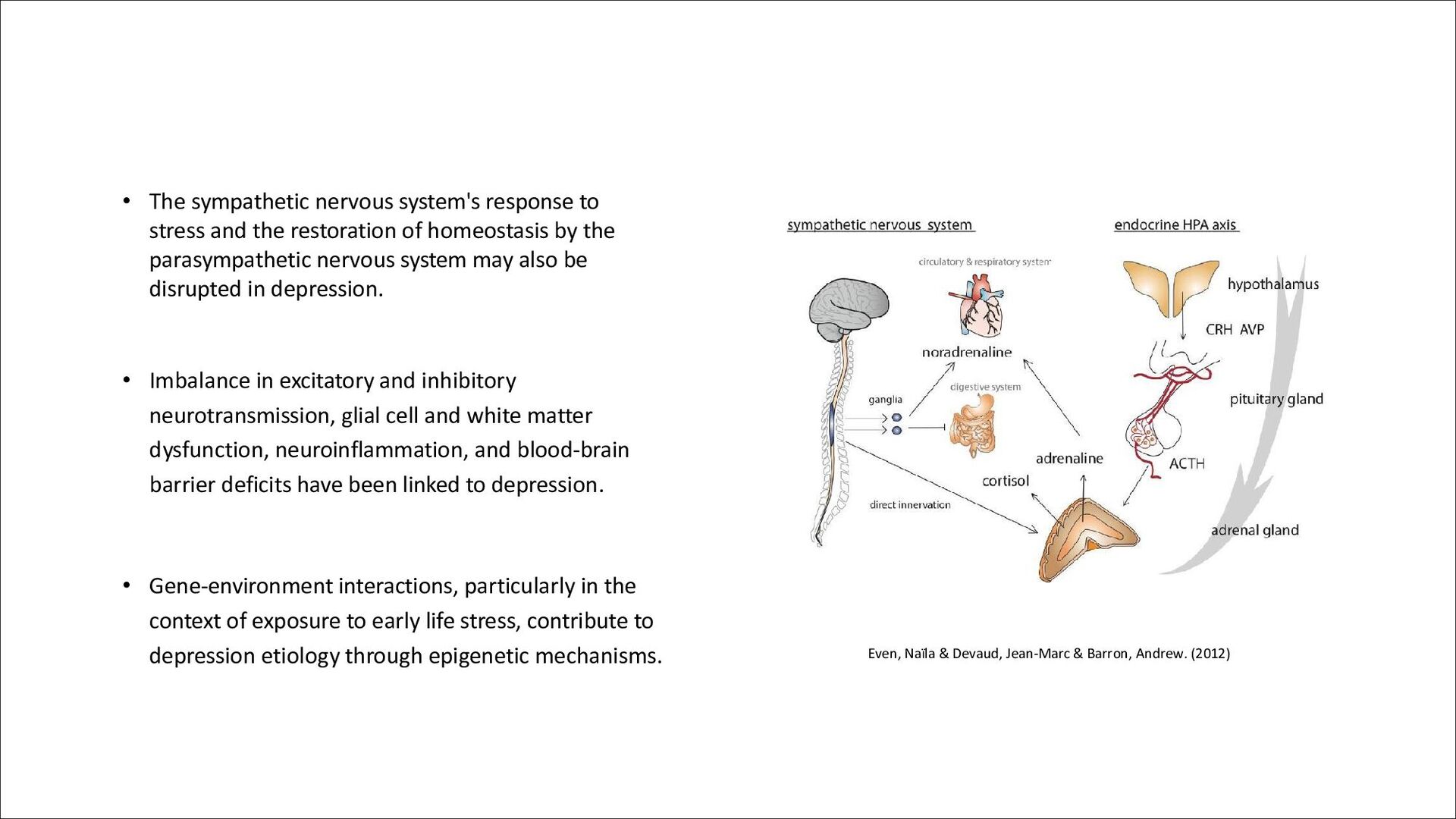
restoration of homeostasis by the parasympathetic nervous system may also be disrupted in depression. • Imbalance in excitatory and inhibitory neurotransmission, glial cell and white matter dysfunction, neuroinflammation, and blood-brain barrier deficits have been linked to depression. • Gene-environment interactions, particularly in the context of exposure to early life stress, contribute to depression etiology through epigenetic mechanisms. Even, Naïla & Devaud, Jean-Marc & Barron, Andrew. (2012)
of MDD, and they are more likely to experience comorbid anxiety and recurrent depression. Men are more likely to experience comorbid substance use disorder and have a higher risk of death by suicide. • Brain imaging studies have revealed sex-specific differences in resting state functional connectivity and structural imaging patterns associated with depression. Some differences are the default mode network and cortical thickness, surface area, and gyrification.
sex-stratified genome-wide association studies identifying distinct loci in men and women. • Sex hormones, as well as differences in neurotransmission systems, neurotrophic factors, and pro- inflammatory and anti-inflammatory cytokines, may contribute to sex differences in depression. • Transcriptomic studies have shown strikingly distinct gene expression changes in males and females with depression (sex-specific molecular alterations)
Paradigms: • Chronic variable stress (CVS), chronic unpredictable mild stress (CUMS), and social isolation paradigms produce different responses in male and female rodents. • Learned helplessness paradigm shows less pronounced sex-specific effects. Early Life Stress Models: • Maternal separation shows effects in both males and females. • Chronic social defeat stress (CSDS) has recently been adapted for use in female mice. Biological Systems of Interest: • Components of the HPA axis, including the GR and CRH, show sex-specific differences in response to stress. • Brain-derived neurotrophic factor (BDNF), serotonin receptors and transporters, and GABA-ergic and glutamatergic signaling components show some evidence of sex differences in animal models of stress.
in resilience and susceptibility in stress models, mediated through components of the HPA axis. Transcriptomic and Behavioral Responses: • Sex-specific changes in response to cortisol administration, including behavior, blood cortisol levels, and adrenal gland weights. • Sex-specific increases in peripheral inflammatory markers in response to CSDS, partially reversed by phytochemical combination therapy. • Transcriptomic effects of stress on the PFC show sex-specific responses, partially rescued by combination therapy of phytochemicals. Microglial States and Immune Function: • Sex differences in microglial states and expression of immune function-related genes at baseline and in response to stress. • Differential contributions of sex chromosomes and gonadal hormones to sex differences in stress response within rodent models.
brain Labonté et al.: • Little overlap of MDD-associated DEGs in males and females. • Lack of concordance in threshold-free patterns of MDD-associated transcriptomic change between the sexes. • Gene co-expression module analysis supported sex differences in MDD-associated gene expression changes. Studies on lincRNAs: • Striking differences in MDD-associated changes in expression of lincRNAs between males and females. • Baseline differences in lincRNA expression in the brain between the sexes.
MDD using a lenient FDR of 25%. • 39 of these genes showed an effect of sex, although not necessarily revealing opposite directions of effect between males and females. Ding et al. (2018): • Most DEGs were sex-specific, and the DEGs in common between the sexes primarily showed opposite directions of effect. • In threshold-free comparisons, the authors observed evidence for discordance in patterns of MDD- associated gene expression between males and females.
ITPR3) with evidence for overall differential expression in MDD. • Found a handful of mutually exclusive DEGs in male (including ITPR3) and female (including SPRY2) specific sex-stratified analysis. • Found 6 genes with opposite direction of effect in males and females when examining sex by phenotype interactions, including CKB and UBE2M which were increased in expression in the depressed males, and MANEA which was increased in expression in the depressed females per sex- stratified analysis.
glial number and morphology in cortical areas, specific to cell subtypes or cortical layers, in MDD. • Transcriptome-wide studies implicated genes related to synaptic vesicles (e.g., SYN1, CALM2), dendritic spines (RAB4B), and axon growth (TUBB4) showed decreased expression in MDD. • Specific transcript variants of SYN1 and SYN2 (synaptic proteins involved in neurotransmitter release and synaptic function) were increased in expression in the PFC in individuals with MDD. • Gene Co-Expression Analysis: Identified a gene module enriched for glutamatergic and GABAergic gene sets, as well as containing genes related to neuron growth and development (e.g., BDNF, ephrins).
glutamatergic and GABAergic neurotransmission systems in the cortex have been reported in depression. • Deficits in the GABAergic neurotransmission system in the cortex have been implicated, strongly involving somatostatin expressing neurons, and to some extent parvalbumin (PV) expressing neurons. • Glutamate receptor gene expression studies found higher expression in cases than controls, with majority of glutamate receptor genes showing increased expression in female cases compared to female controls specifically.
functions were downregulated in depressed individuals, including those encoding connexins (GJA1, GJB6) and the SOX9 transcription factor. • Astrocyte Morphology: Changes in morphology, with white matter astrocytes showing more elaborate branching patterns in the ACC in individuals with depression.
function, such as MAG, MOG, MOBP, and PLP1, were downregulated in MDD. • Gap Junctions Dysregulation: decreased expression of connexin genes (GJB1 and GJC2) and dysregulation of gap junctions between astrocytes and oligodendrocytes (OCLDN, CAV1, CAV2), were reported. Microglia and Endothelial Cells • Dysregulation of inflammatory molecules including cytokines and molecules downstream of interferons in MDD. • Studies found an increased proportion of primed to ramified microglia in depressed individuals, and changes in endothelial cell gene expression, suggesting altered blood brain barrier and endothelial cell molecular state in MDD.
and genome sequencing in post-mortem human brain tissue. This has given us unprecedented access to cell-type specific gene expression profiles and somatic mutations unique to pathological states. Additionally, we can now measure epigenetic information from individual brain cells and with advanced statistical approaches, we can focus on the key cell-types underlying psychiatric and other brain phenotypes”. Sequencing the human brain at single-cell resolution
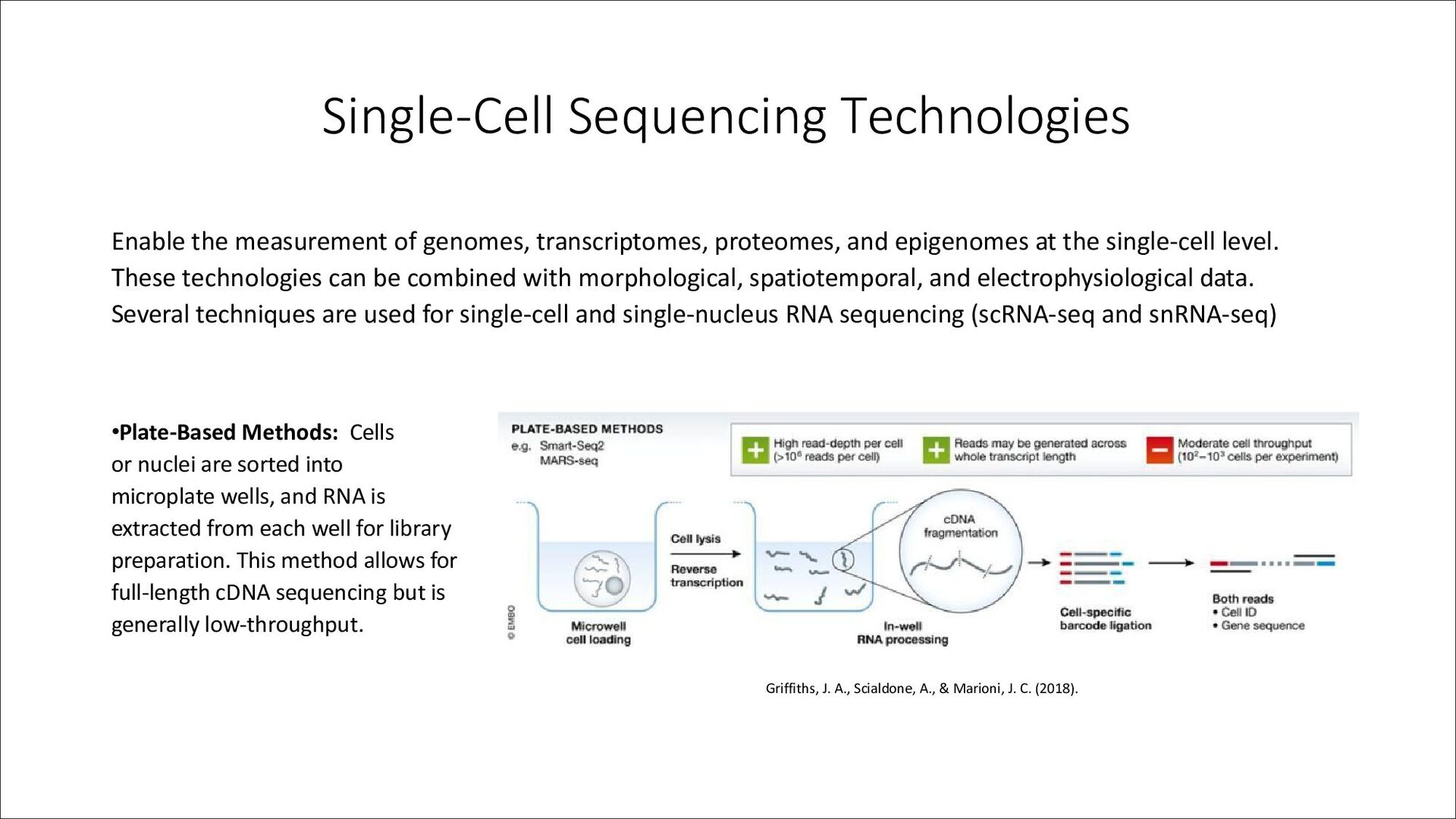
and epigenomes at the single-cell level. These technologies can be combined with morphological, spatiotemporal, and electrophysiological data. Several techniques are used for single-cell and single-nucleus RNA sequencing (scRNA-seq and snRNA-seq) •Plate-Based Methods: Cells or nuclei are sorted into microplate wells, and RNA is extracted from each well for library preparation. This method allows for full-length cDNA sequencing but is generally low-throughput. Griffiths, J. A., Scialdone, A., & Marioni, J. C. (2018).
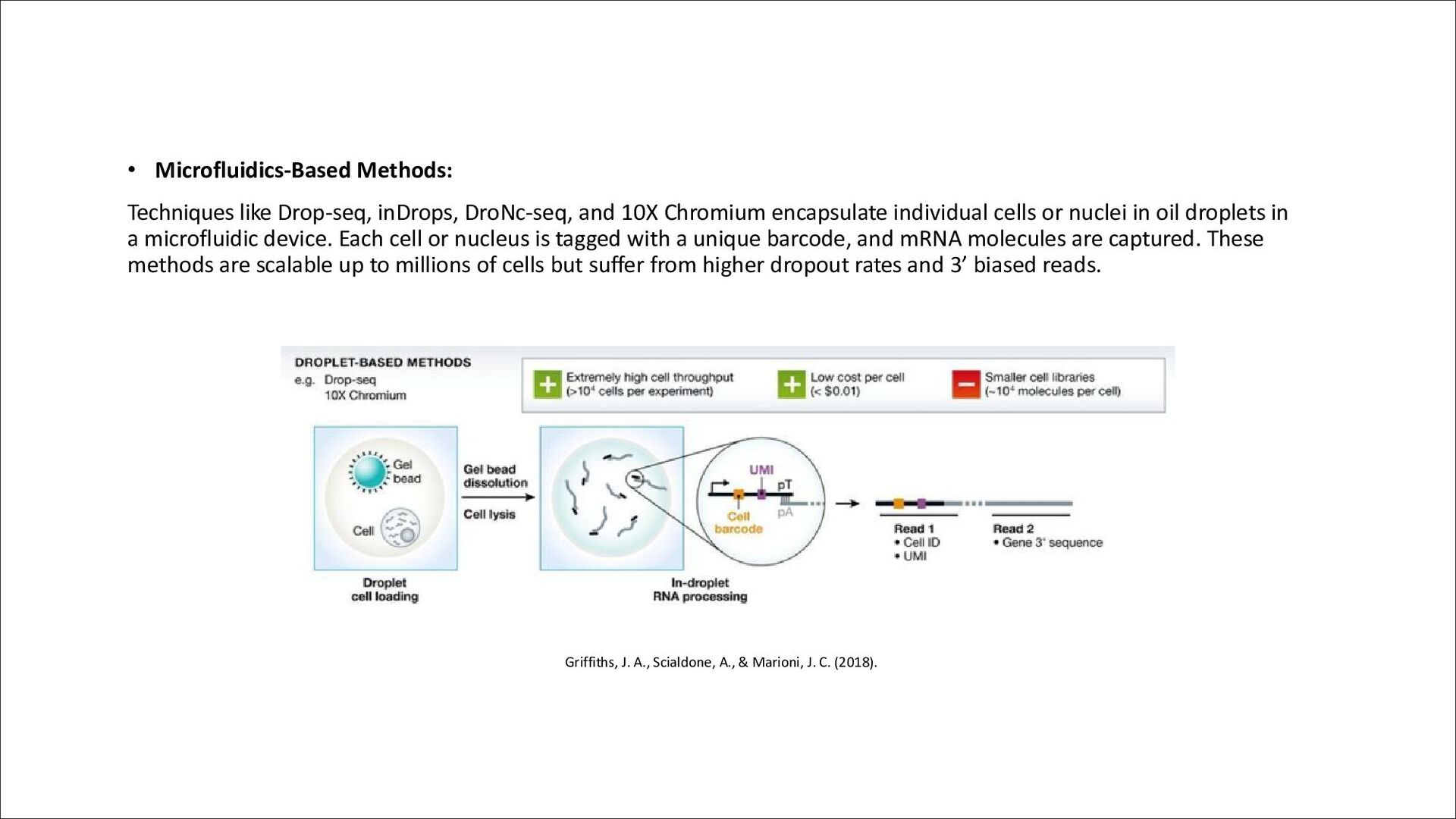
Chromium encapsulate individual cells or nuclei in oil droplets in a microfluidic device. Each cell or nucleus is tagged with a unique barcode, and mRNA molecules are captured. These methods are scalable up to millions of cells but suffer from higher dropout rates and 3’ biased reads. Griffiths, J. A., Scialdone, A., & Marioni, J. C. (2018).
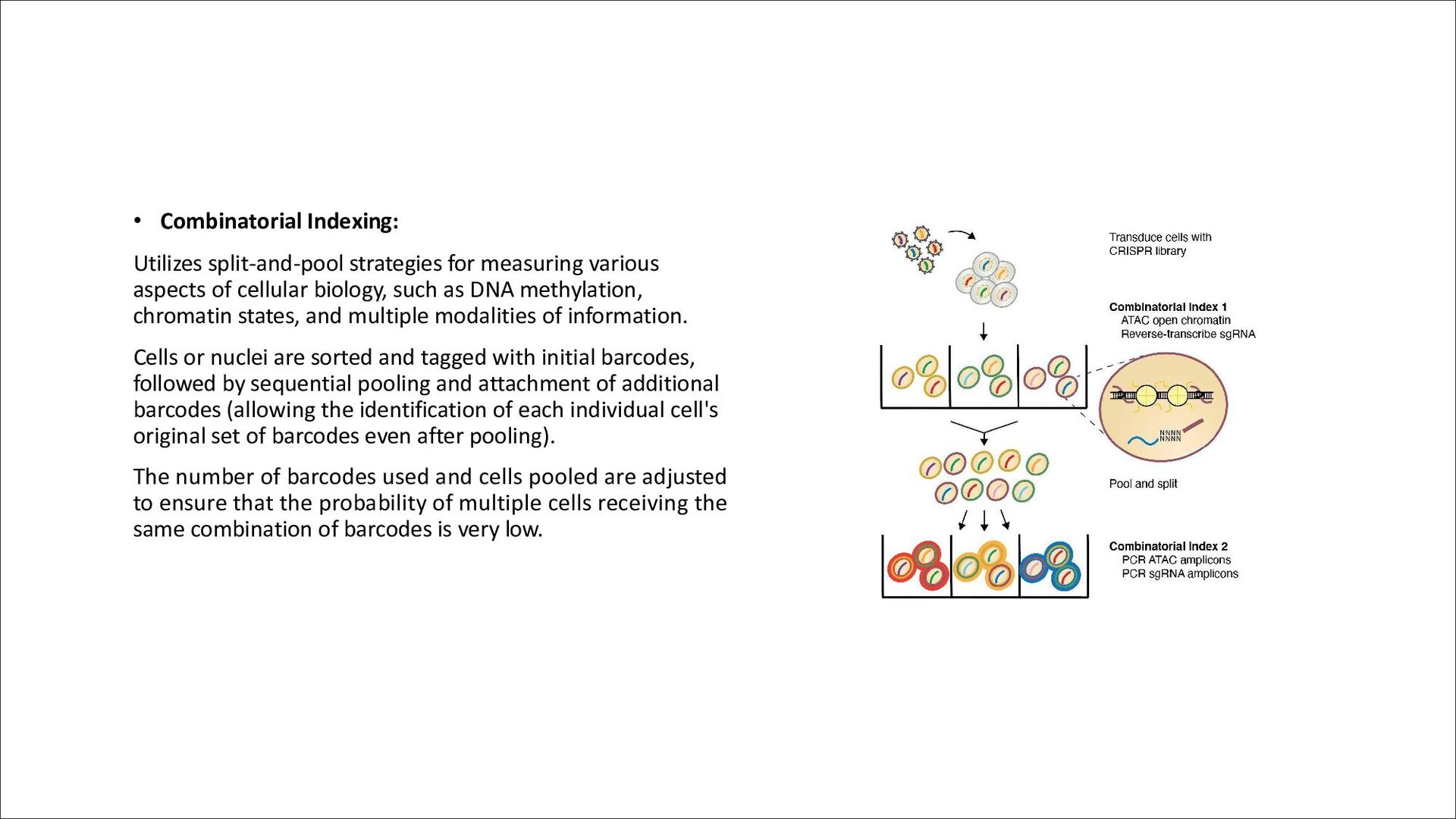
of cellular biology, such as DNA methylation, chromatin states, and multiple modalities of information. Cells or nuclei are sorted and tagged with initial barcodes, followed by sequential pooling and attachment of additional barcodes (allowing the identification of each individual cell's original set of barcodes even after pooling). The number of barcodes used and cells pooled are adjusted to ensure that the probability of multiple cells receiving the same combination of barcodes is very low.
most studies thus far have relied on sorting of individual nuclei into the wells of a microplate. However, recently, commercial technology for droplet-based high-throughput single-nucleus genome sequencing has been created which holds promise for detecting both copy-number variations (CNVs) and single-nucleotide variations (SNVs) at single-cell resolution • Droplet-based profiling of open chromatin in single-cells has also been developed. Furthermore, several techniques for acquiring multimodal data at single-cell resolution exist: • CITE-seq: measures cell-surface epitopes and transcriptomes • Patch-seq: measures electrophysiological recordings and transcriptomes
Regions Studied • Cortical Regions: Several studies have profiled cortical regions, revealing diverse cell populations and their gene expression profiles. • Subcortical Structures: Studies have also focused on subcortical structures, providing insights into the cellular composition and function of these regions. • Hippocampus: The hippocampus, known for its role in memory and learning, has been extensively studied at the single-cell level, uncovering distinct cell types and their molecular signatures. Developmental Stages over time • Different stages of brain development have been investigated, shedding light on how cellular diversity and gene expression change over time.
or microglia have been targeted, enabling the characterization of these cells in detail. Experimental Perturbations • Cell-type-specific signatures of various experimental perturbations have been measured, providing insights into how different stimuli affect gene expression in specific cell populations. Cellular Diversity Atlases • Single-cell transcriptomics has led to the creation of large-scale atlases of mouse brain cellular diversity, providing comprehensive catalogs of cell types and their gene expression profiles.
have provided insights into somatic mutations, including single nucleotide variations (SNVs) and copy number variations (CNVs), which may accumulate in post-mitotic cells of the central nervous system over development and aging. CNVs in Single Neurons: • McConnell et al. (2013) studied CNVs in single neurons from post-mortem human frontal cortex and induced pluripotent stem cell (iPSC) derived neurons. They found that cultured neurons had a higher incidence of CNVs compared to neural progenitor cells (NPCs) or fibroblasts, suggesting that somatic mutations may play a role in neuronal development. Single-cell resolution studies of sequence variation in the human brain
CNVs in post-mortem brain tissue and found that aneuploidy is rare but sub- chromosomal CNVs are common. They identified CNVs shared by multiple neurons, indicating that these mutations are not artifacts and may have functional significance. L1 Retrotransposon Insertions • Evrony et al. (2012) examined L1 retrotransposition rates in neuronal nuclei and found that somatic L1 insertions are generally rare, with less than one insertion per neuron on average. However, Upton et al. (2015) reported higher rates of somatic L1 insertions in hippocampal neurons and glia, which was later reanalyzed to be closer to the earlier findings of low insertion rates.
the cortex and identified somatic SNVs, with some mutations likely occurring post-mitotically. They traced the developmental lineage of neurons and found that some mutations were present in non-brain tissues, indicating early developmental origin. Accumulation of SNVs with Age • Lodato et al. (2018) measured SNVs in the hippocampus and prefrontal cortex of individuals spanning different ages. They found that somatic SNVs accumulate with age and are enriched in neuronally expressed genes, with a higher rate of accumulation in the hippocampus. Comparison with Non-Neuronal Cells • A recent study by Chronister et al. (2019) found that neurons harbor more CNVs than non-neuronal or non-neural cells, and these CNVs tend to affect a larger portion of the genomes. They also found a decreased prevalence of neurons with CNVs in their genomes with age, suggesting a potential susceptibility to aging-related loss.
is challenging due to the loss of material during bisulphite conversion and the need for high sequencing coverage for each cell, making it expensive. • Luo et al. performed single-nucleus WGBS on the human frontal cortex, creating a map of DNA methylation at single- cell resolution. They used fluorescence-assisted nuclei sorting (FAN) to isolate nuclei into microplates, followed by bisulphite conversion and sequencing. -> Map of DNA methylation at single-cell resolution on the human frontal cortex • Analyzed almost 3000 nuclei from the frontal cortex of a single subject. Despite data sparsity, they were able to separate cortical excitatory and inhibitory neuronal subtypes based on DNA methylation signatures, comparable to single-nucleus RNA sequencing (snRNA-seq). • Found that non-CG methylation was more cell-type specific than CG methylation, and patterns of cell-type-specific methylation were highly conserved from mouse brain to human brain. • Valuable reference for cell-type specific DNA methylation in the human brain Single-nucleus methylomics in the post-mortem human brain
insights into cellular diversity, allowing for the identification and characterization of different cell types, including neurons and glial cells. • These studies have revealed differences in gene expression, DNA methylation, and other molecular features between cell types, providing a deeper understanding of brain function and development. • Computational methods, such as imputation and dataset alignment algorithms, have been used to overcome some limitations of single-cell sequencing, such as high gene dropout rates and inter-individual variability. Insights
include underrepresentation of glial cells and lower RNA molecule detection in glial cells compared to neurons. • Technical variability in single-cell sequencing experiments (different samples or individuals). One strategy to address this challenge is to introduce controlled variability during the nuclei capture step (Ex. combine subjects that differ in known SNVs) • Computational challenges, such as analyzing and integrating multimodal data, remain a hurdle in fully harnessing the potential of single-cell sequencing. Challenges
our understanding of disease states by elucidating cell-type contributions to observed disease-related changes in gene expression or DNA methylation in bulk tissue studies. • Integration of multimodal data and use of complementary approaches, such as high-throughput in situ hybridization (ISH), are likely to be key in future single-cell resolution studies of the human brain. • Continued advancements in single-cell sequencing technologies and computational methods will further enhance our ability to study cellular diversity and function in the human brain. Perspectives
of the dorsolateral prefrontal cortex in depression. • Subermaniam, K., Teoh, S. L., Yow, Y. Y., Tang, Y. Q., Lim, L. W., & Wong, K. H. (2021). Marine algae as emerging therapeutic alternatives for depression: A review. Iranian journal of basic medical sciences, 24(8), 997. • Even, N., Devaud, J. M., & Barron, A. B. (2012). General stress responses in the honey bee. Insects, 3(4), 1271-1298. • Griffiths, J. A., Scialdone, A., & Marioni, J. C. (2018). Using single-cell genomics to understand developmental processes and cell fate decisions. Molecular systems biology, 14(4), e8046.
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}