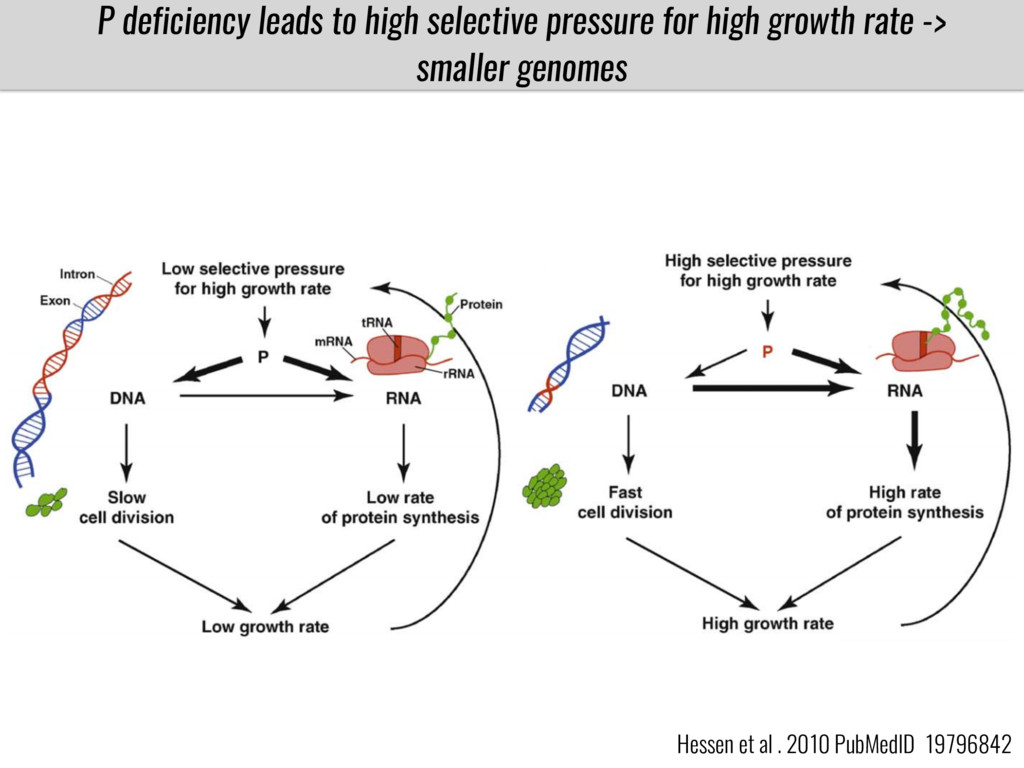
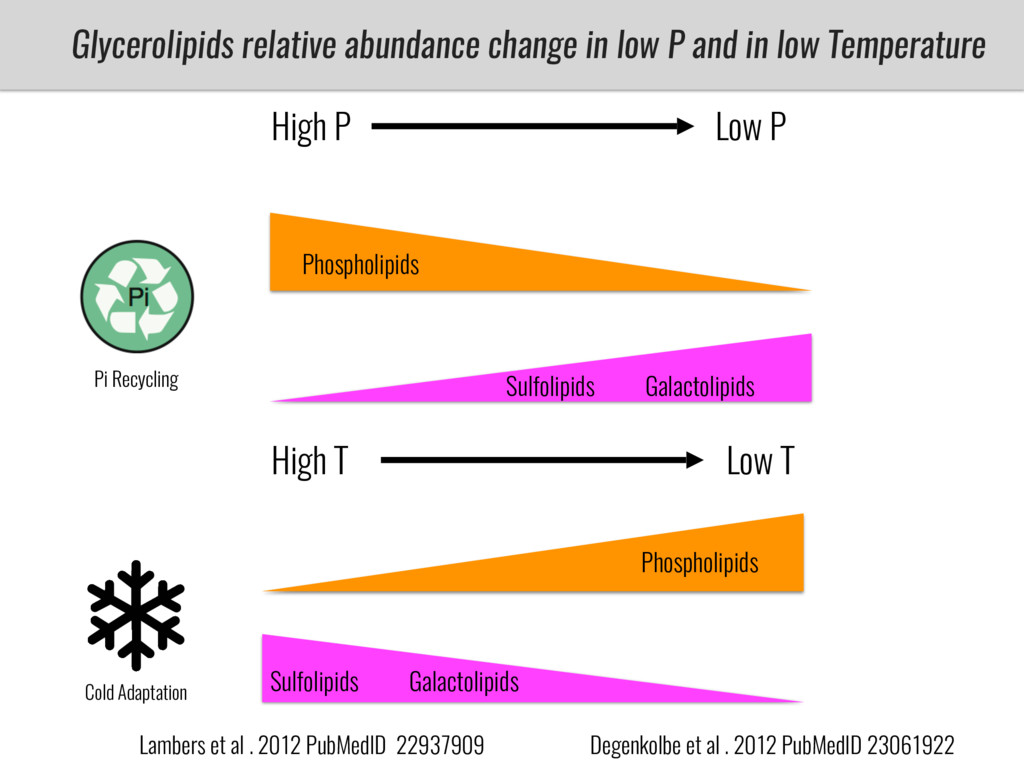
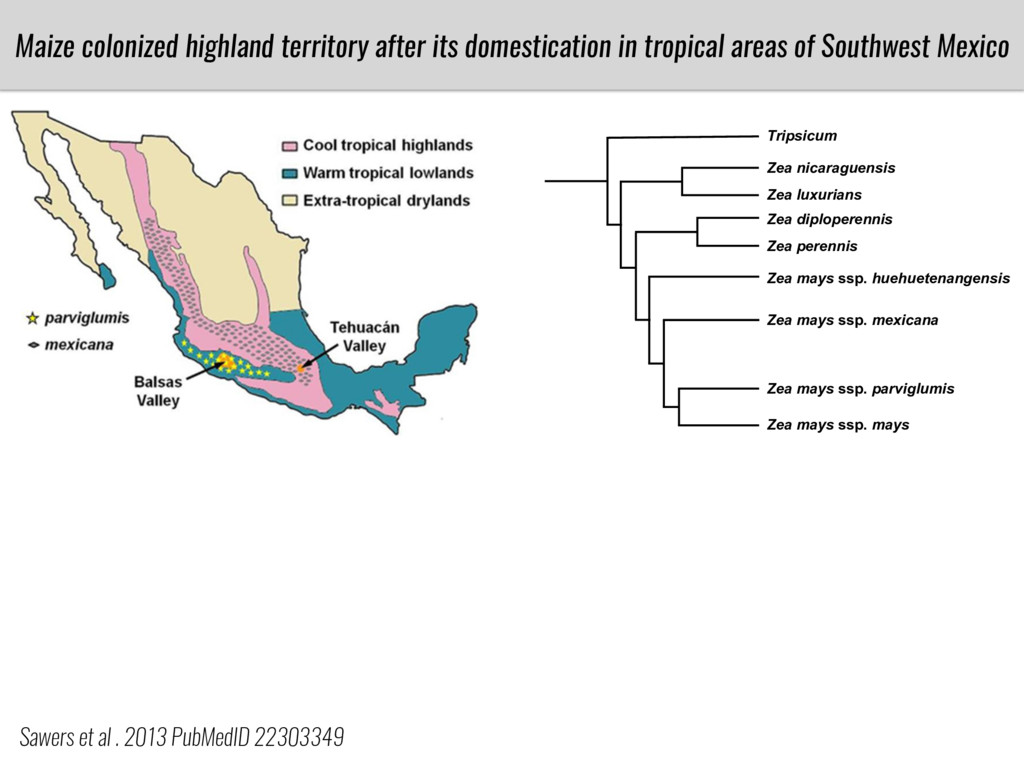
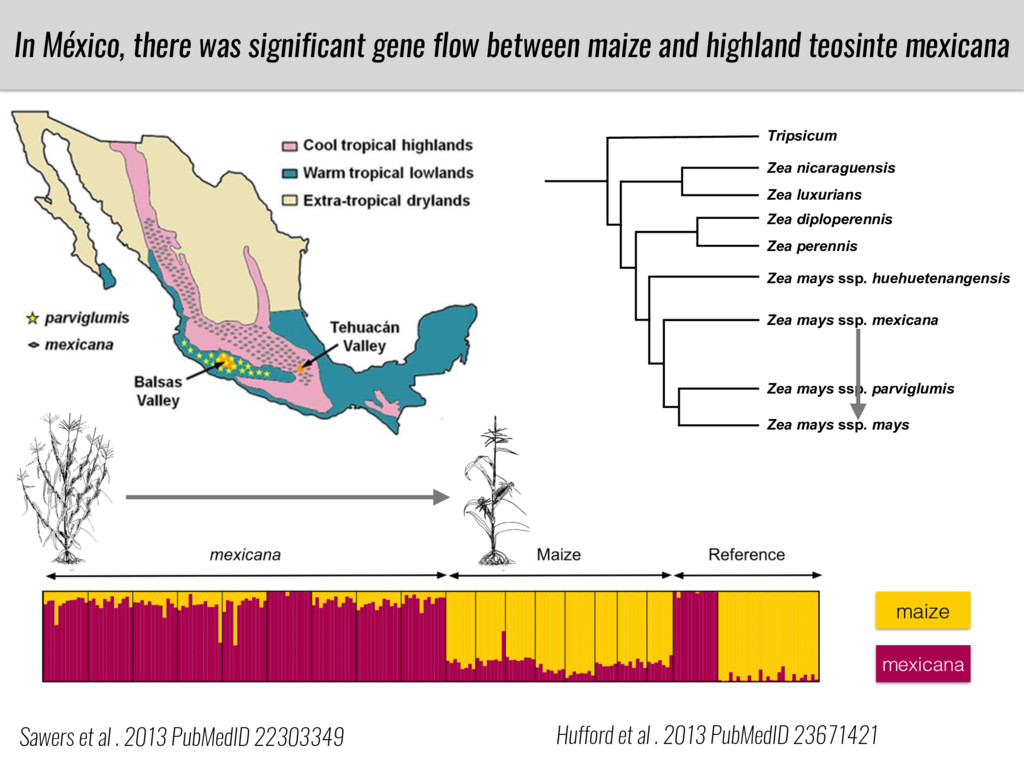
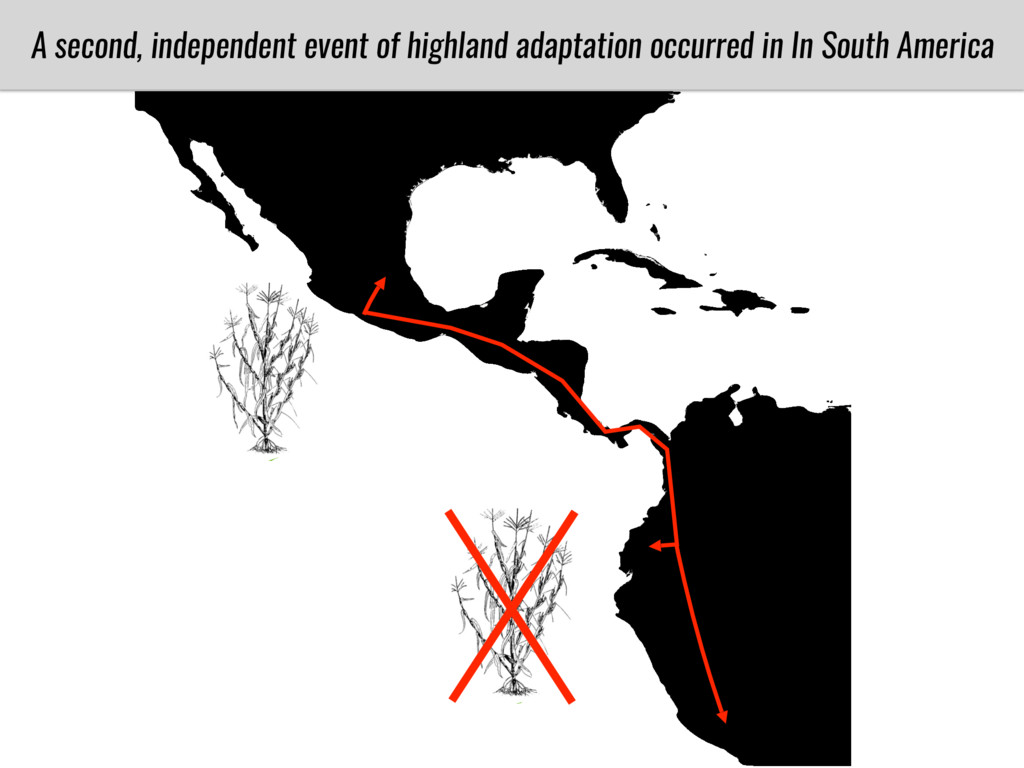
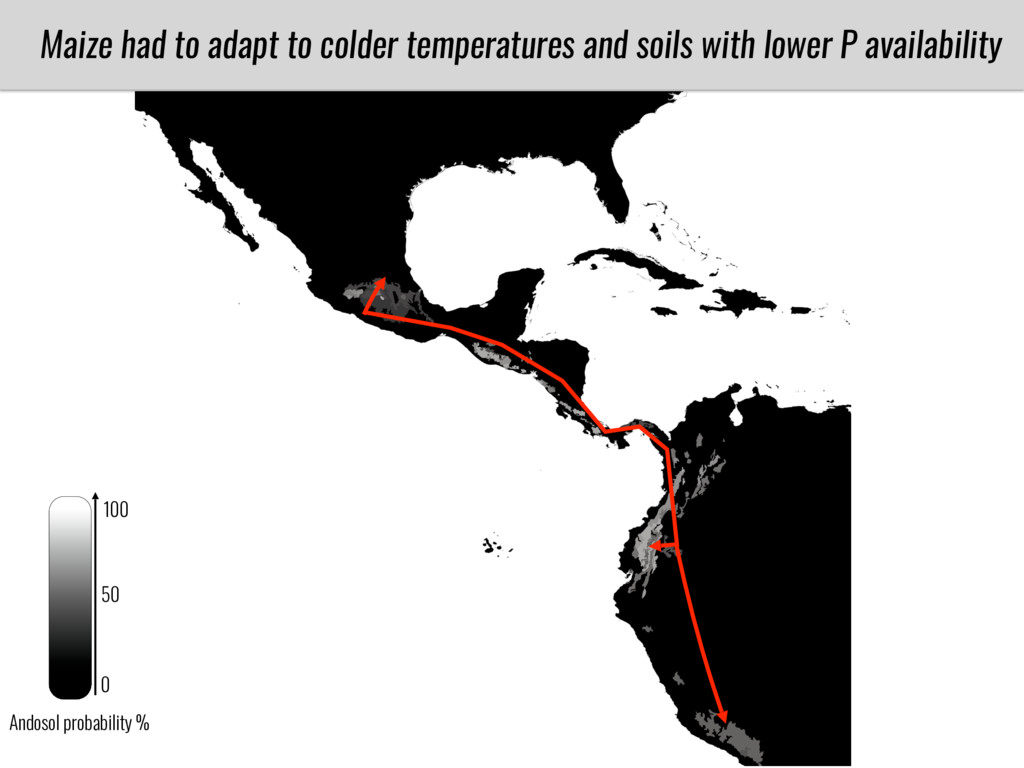
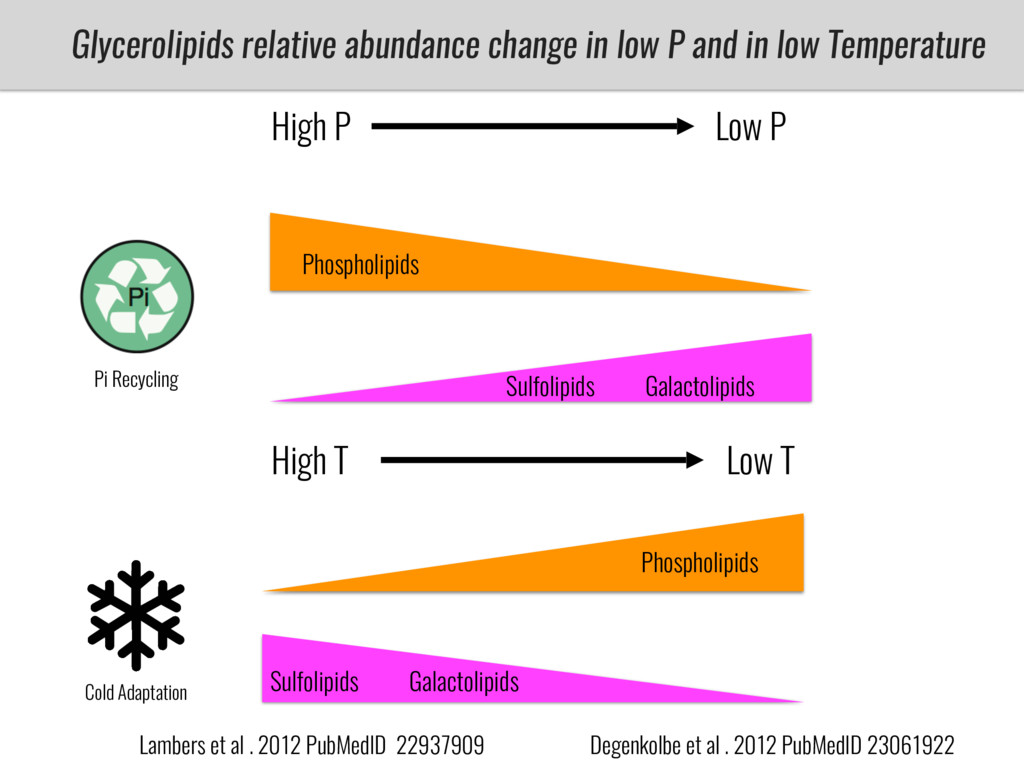
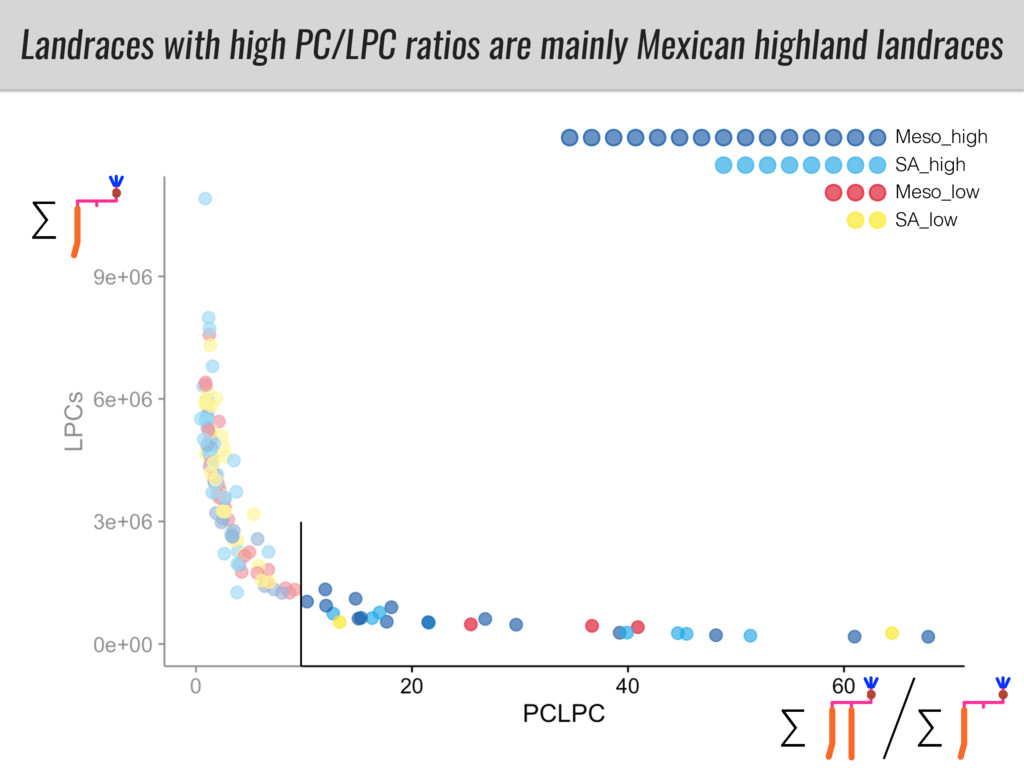
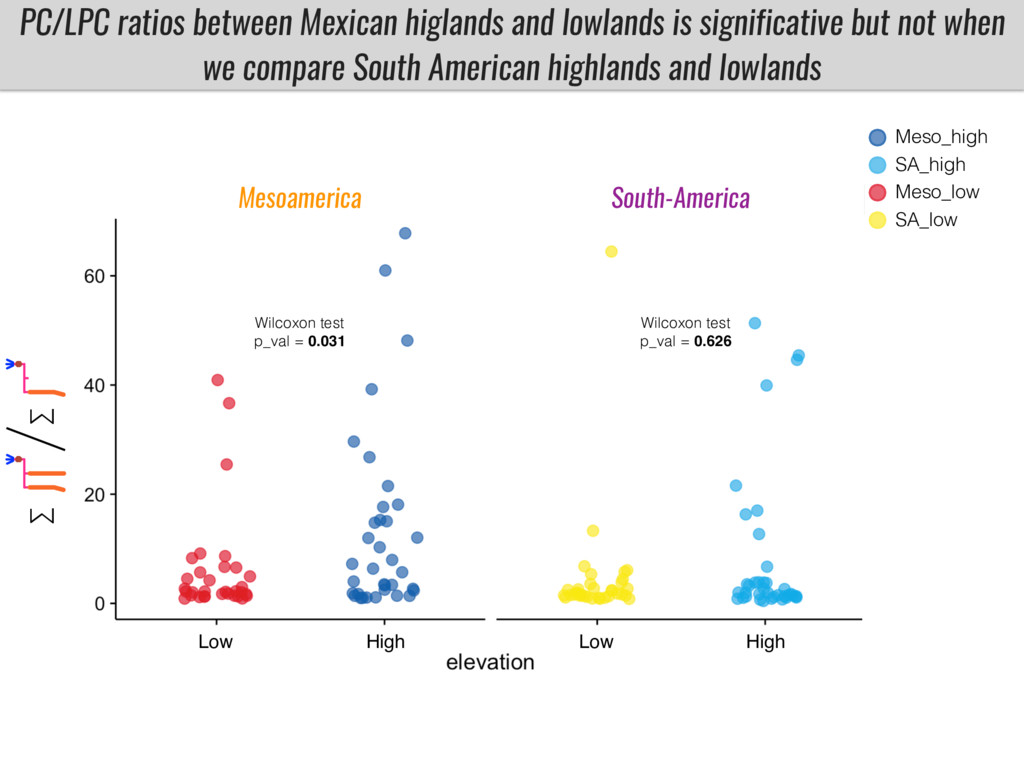
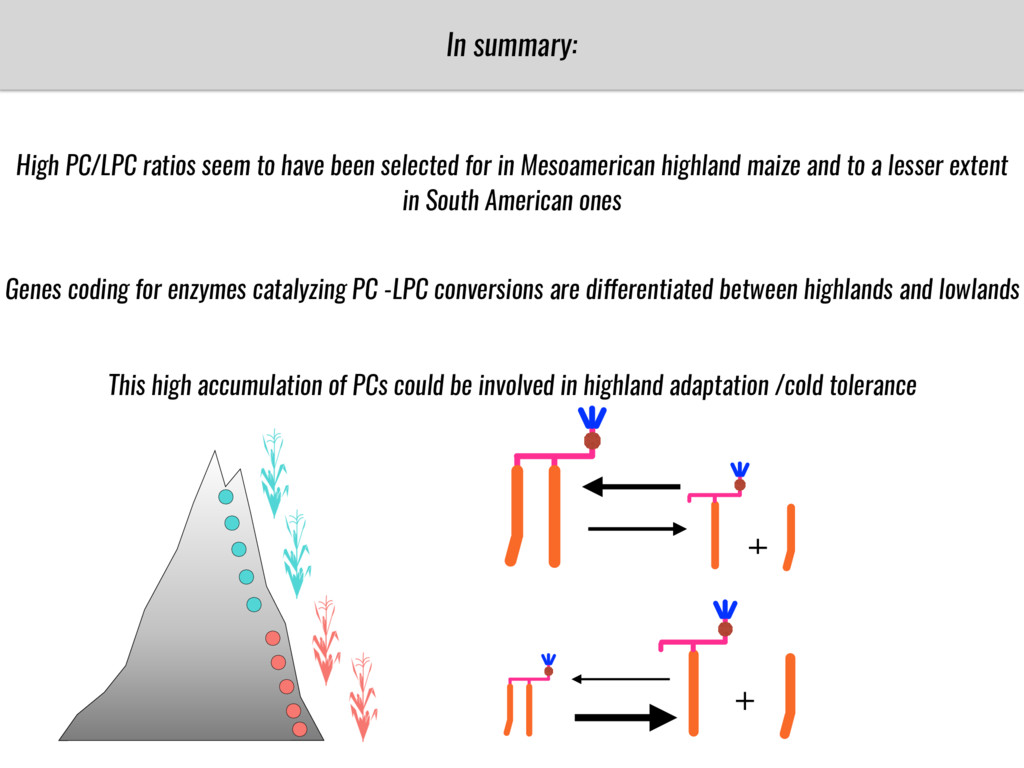
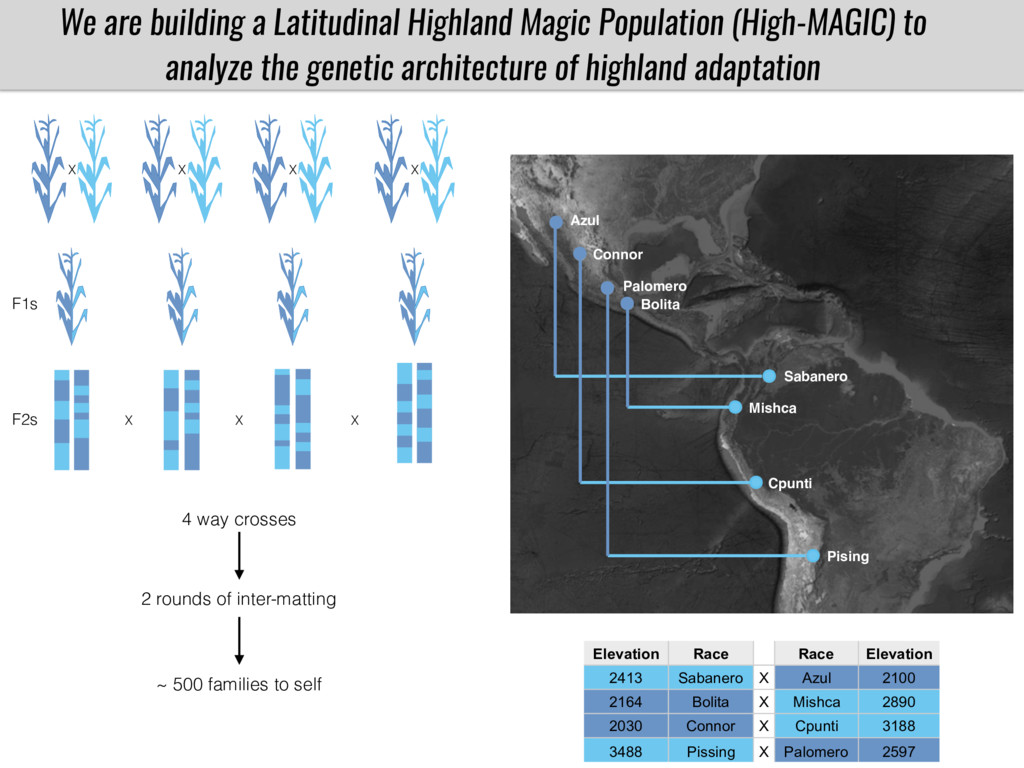
After domestication from lowland teosinte in the Mexican subtropics maize colonized the highlands of Mexico and South America. In both Mexican and South American highlands maize encountered lower temperatures and in several volcanic area soils with low phosphorus availability. We hypothesize that these two abiotic stresses were major selective forces during maize adaptation to the highlands and shaped glycerolipid metabolism since low temperature and P deficiency have opposite effects on the relative content of glycerolipid species. We are using this system to explore how metabolic pathways change during the process of local adaptation and how we can use metabolic phenotyping together with quantitative and population genetics to identify loci that might have been under selection during the local adaptation events.
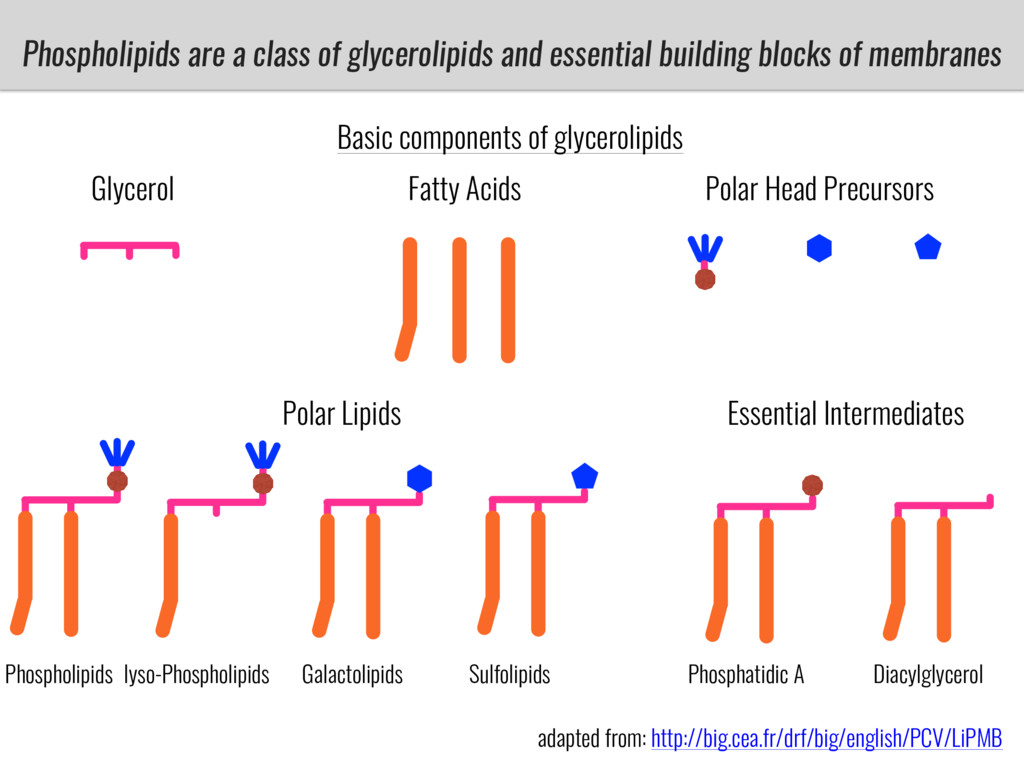
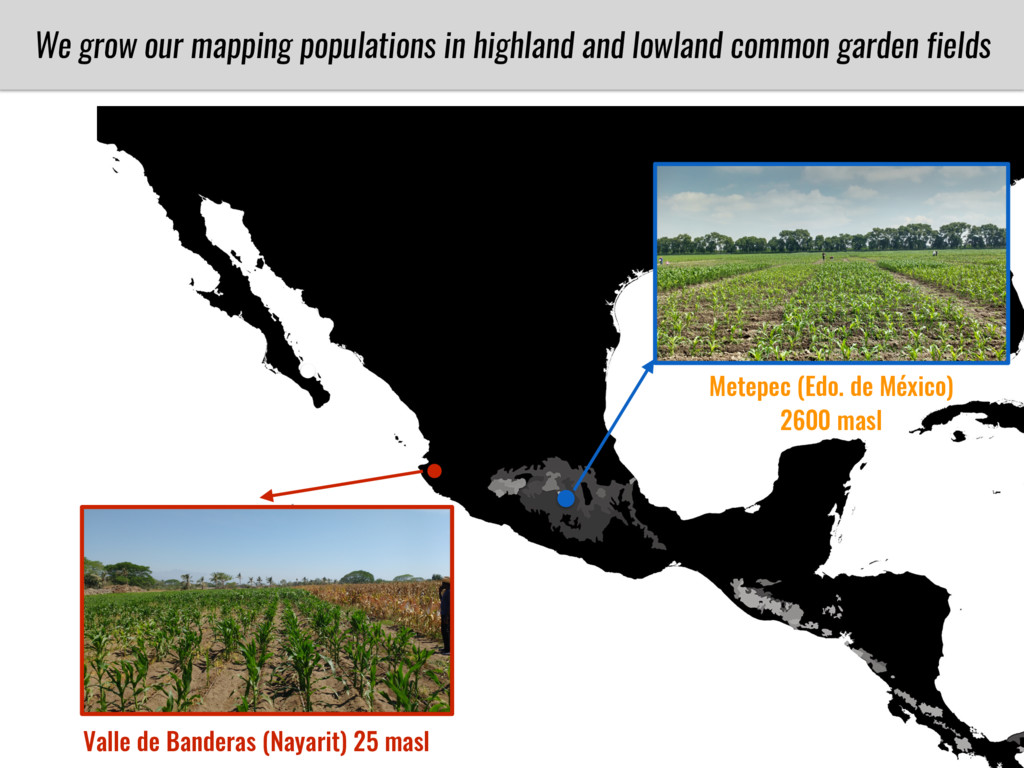
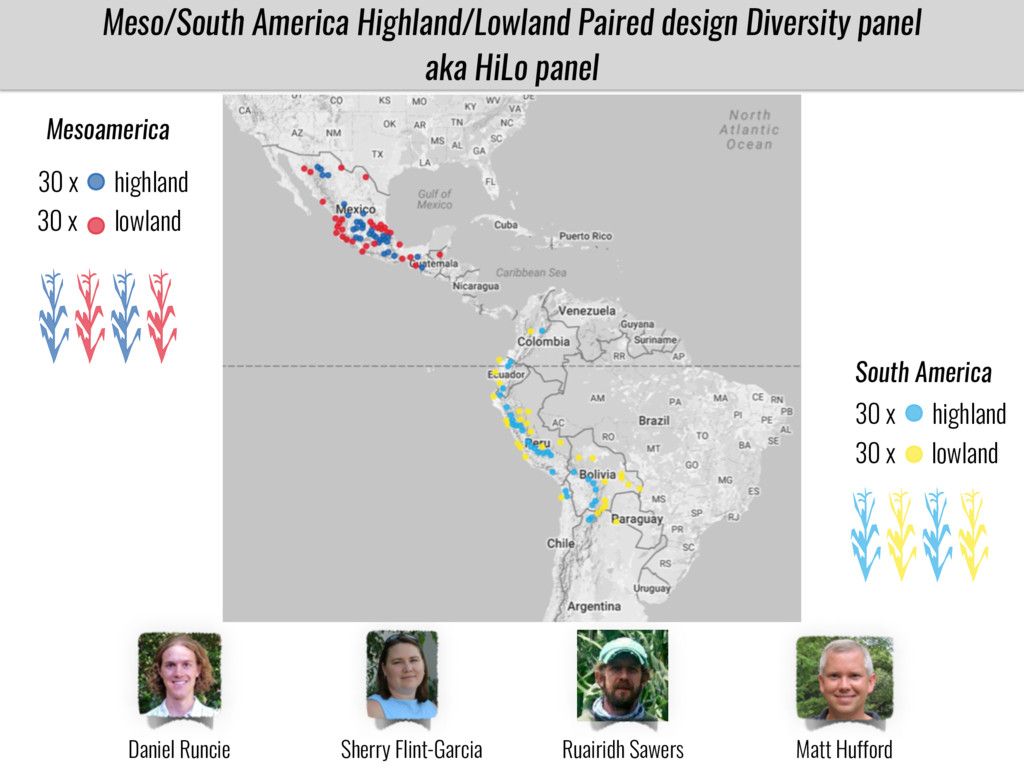
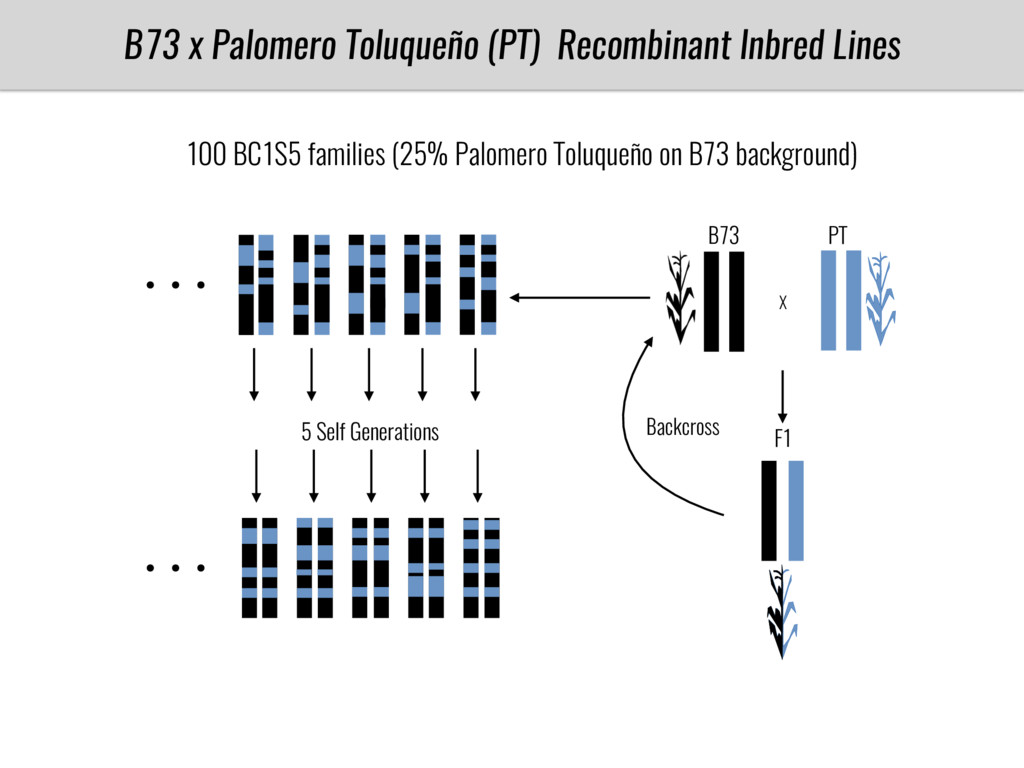
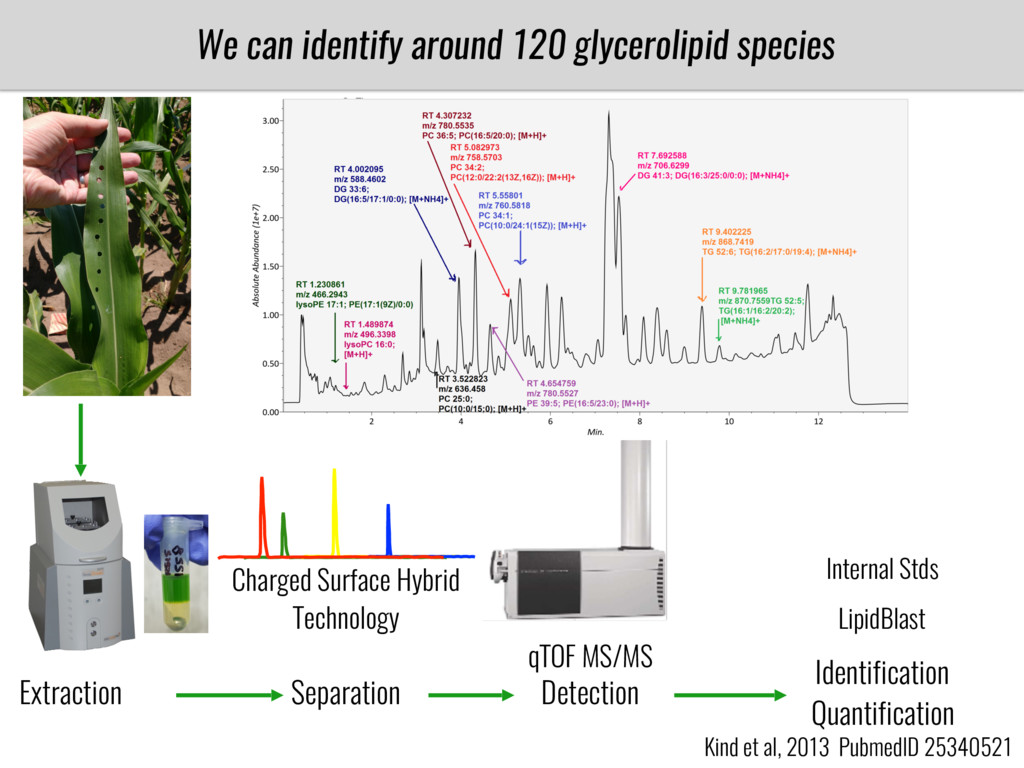
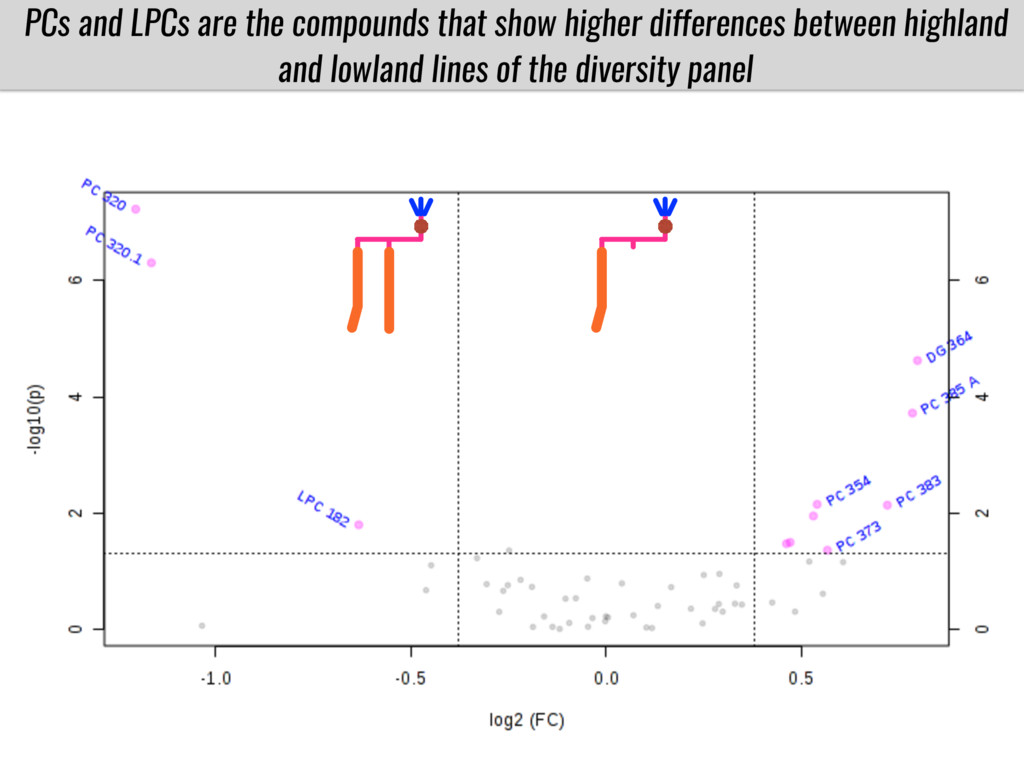
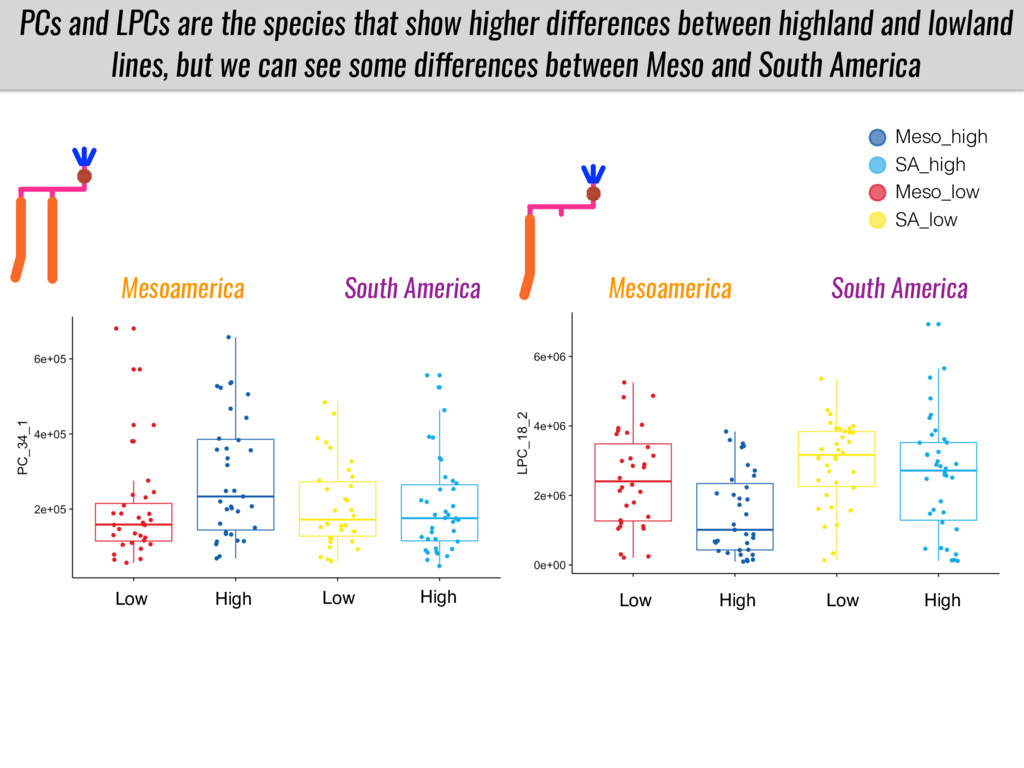
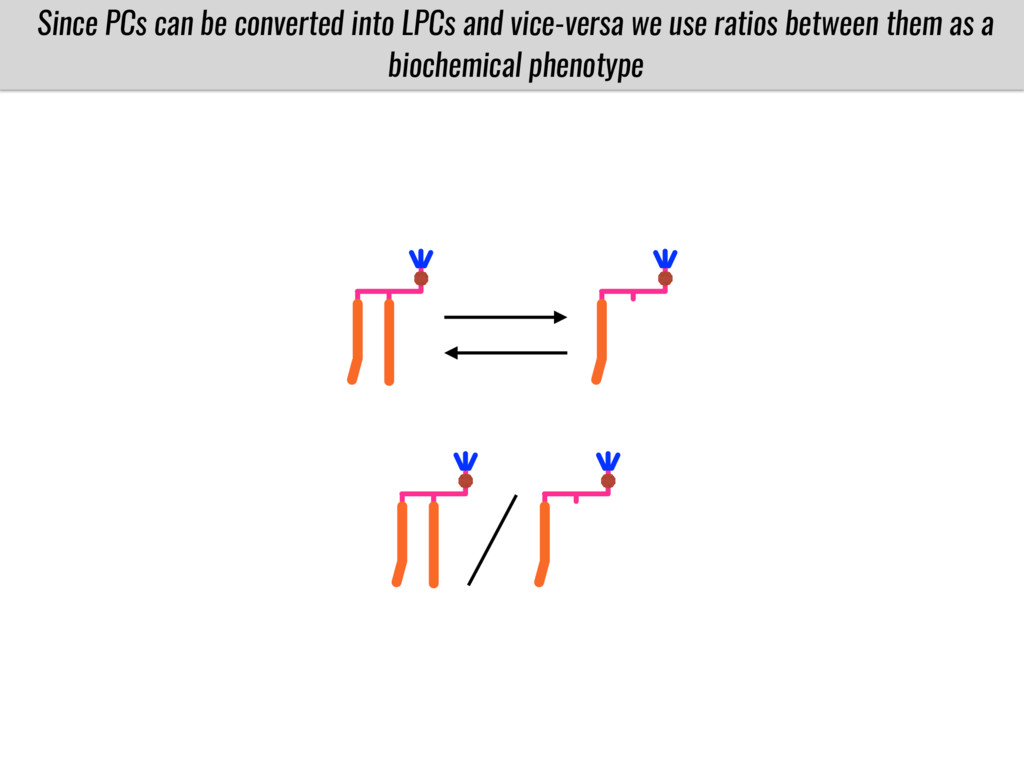
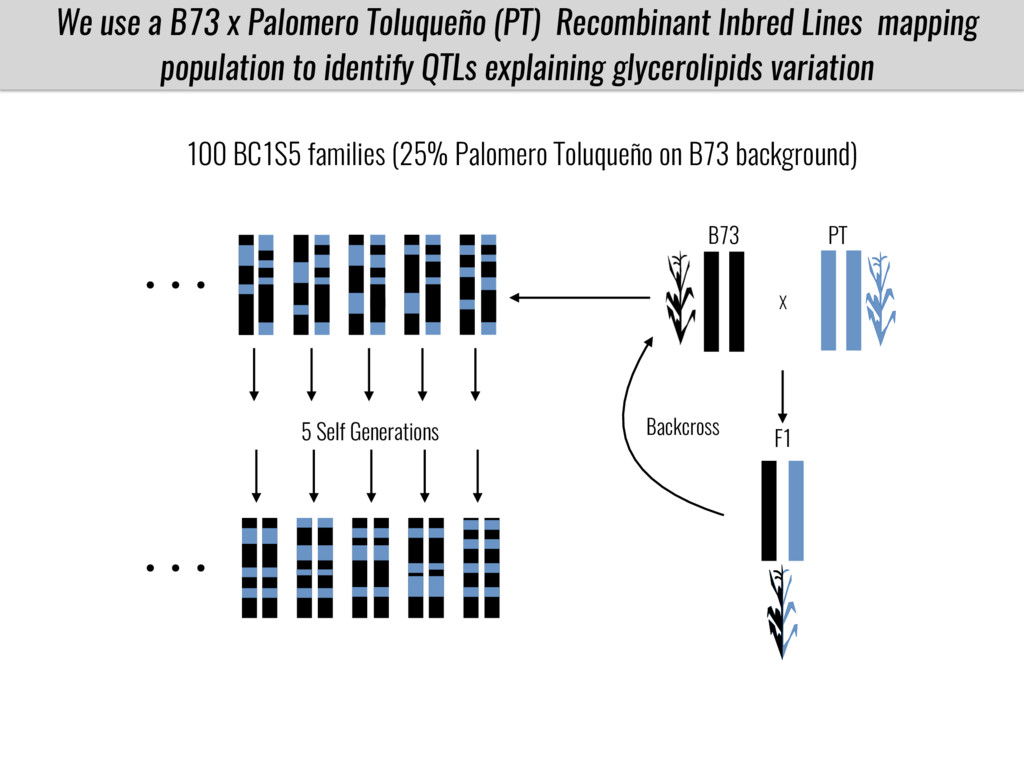
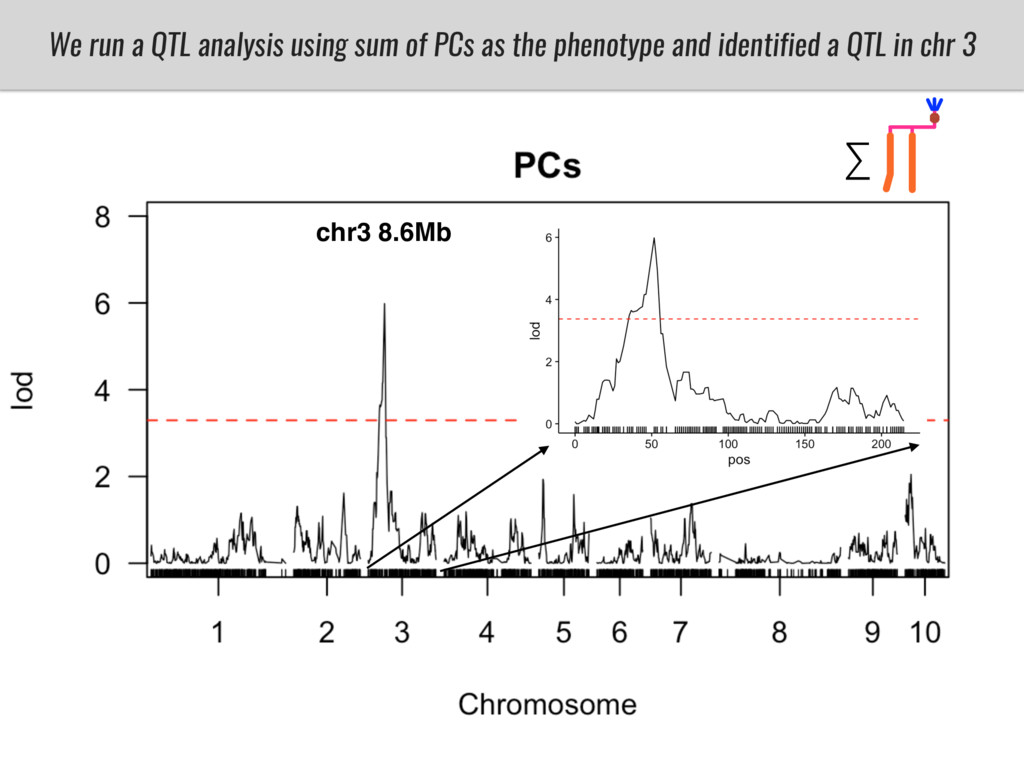
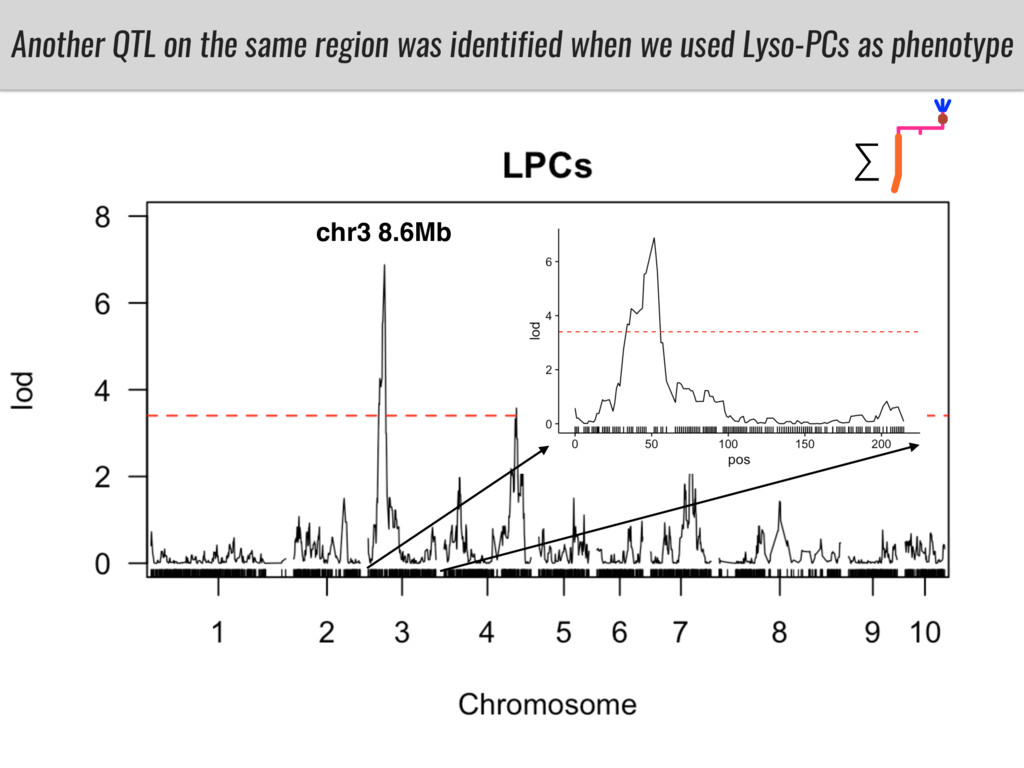
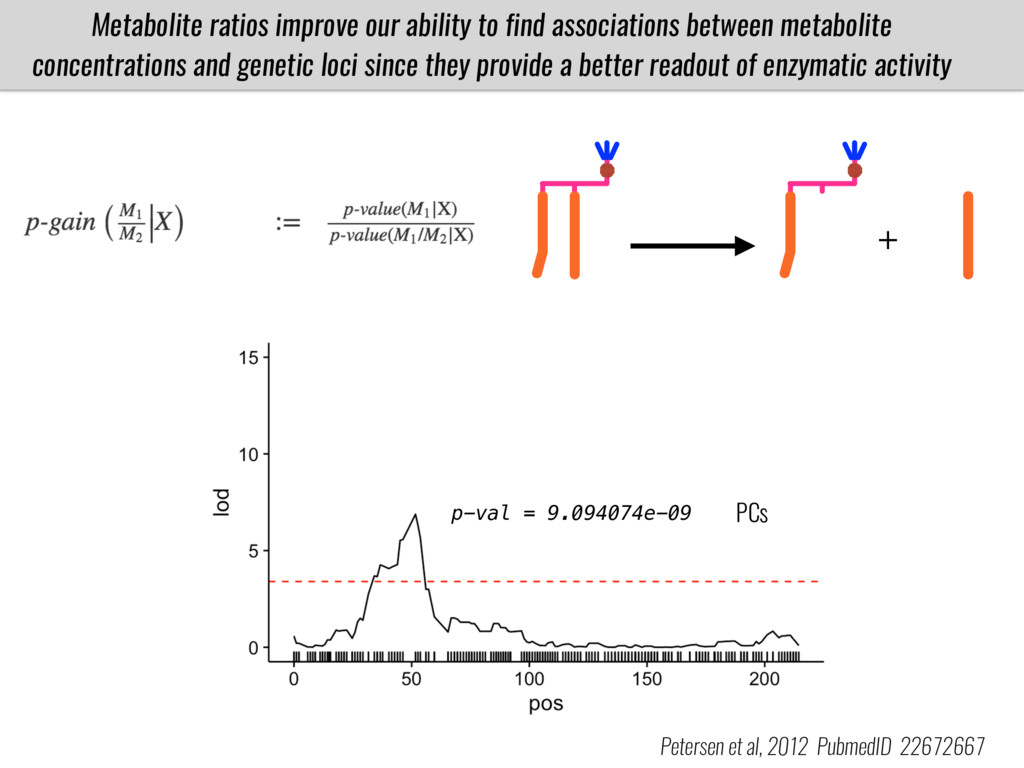
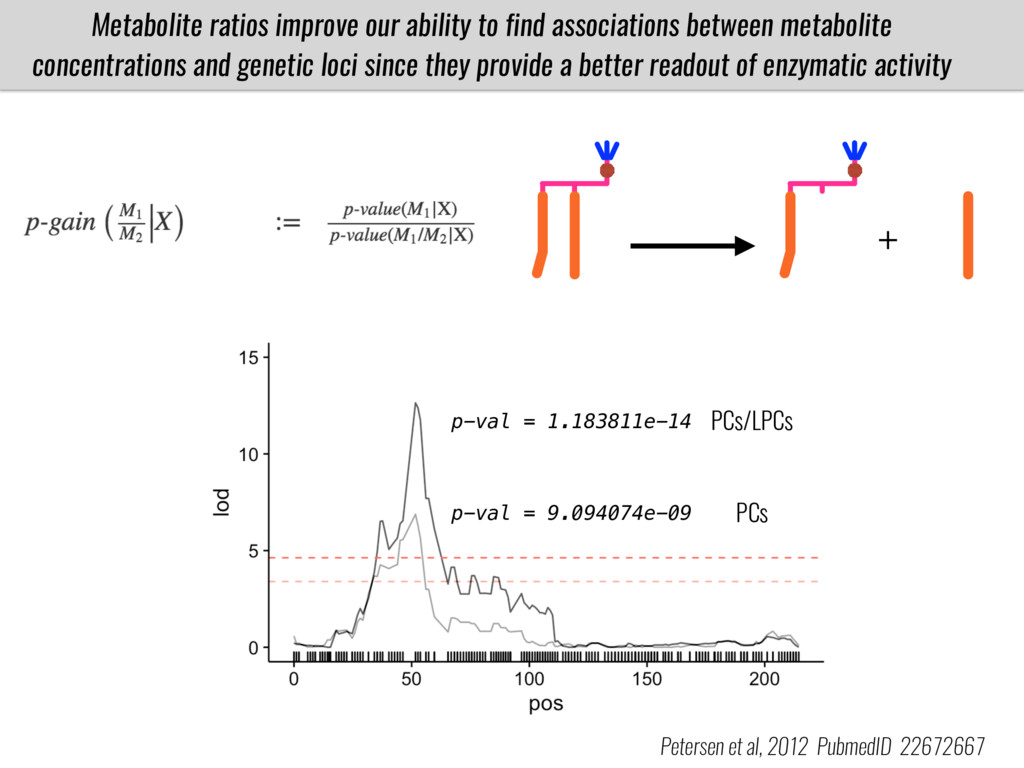
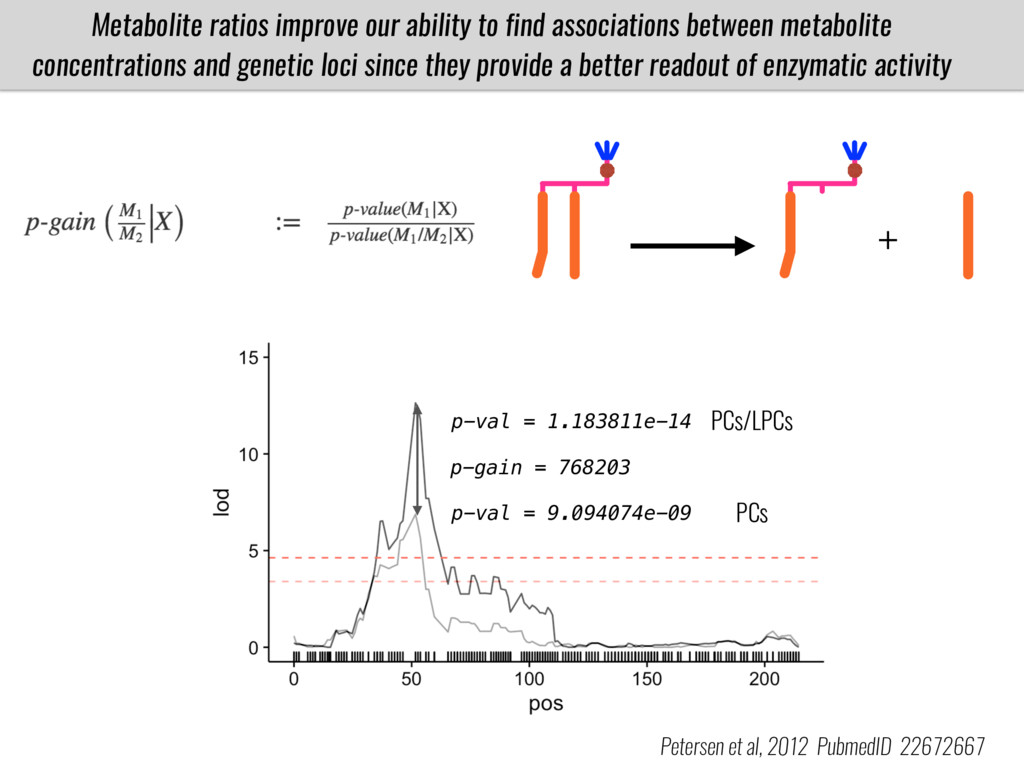
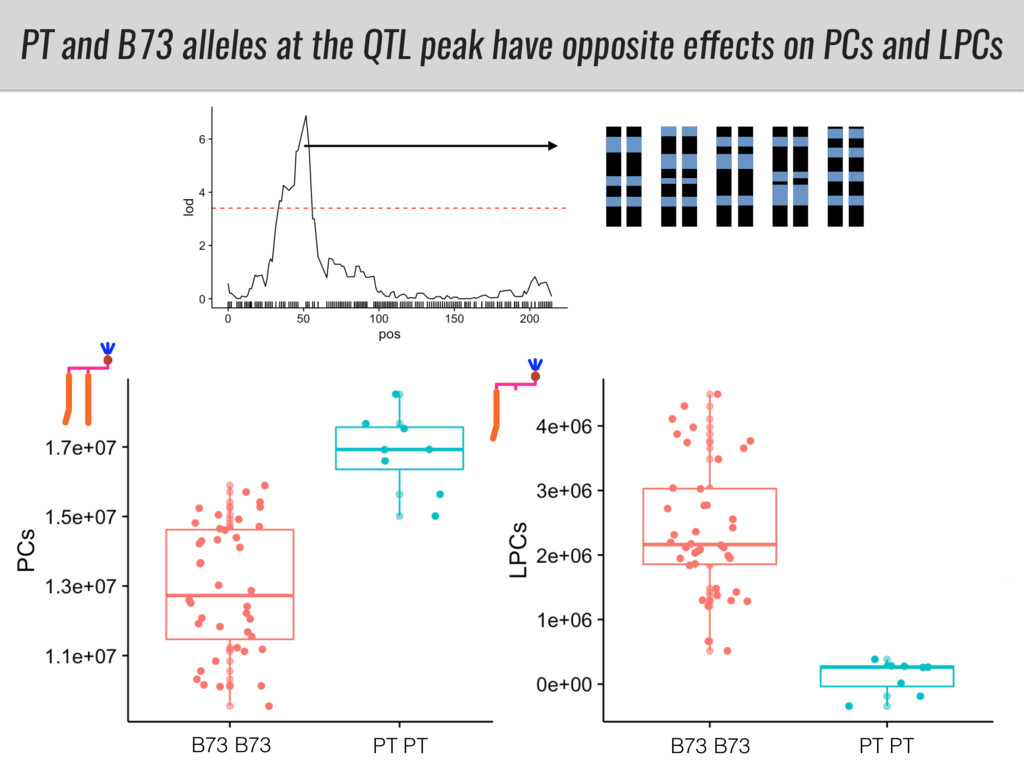
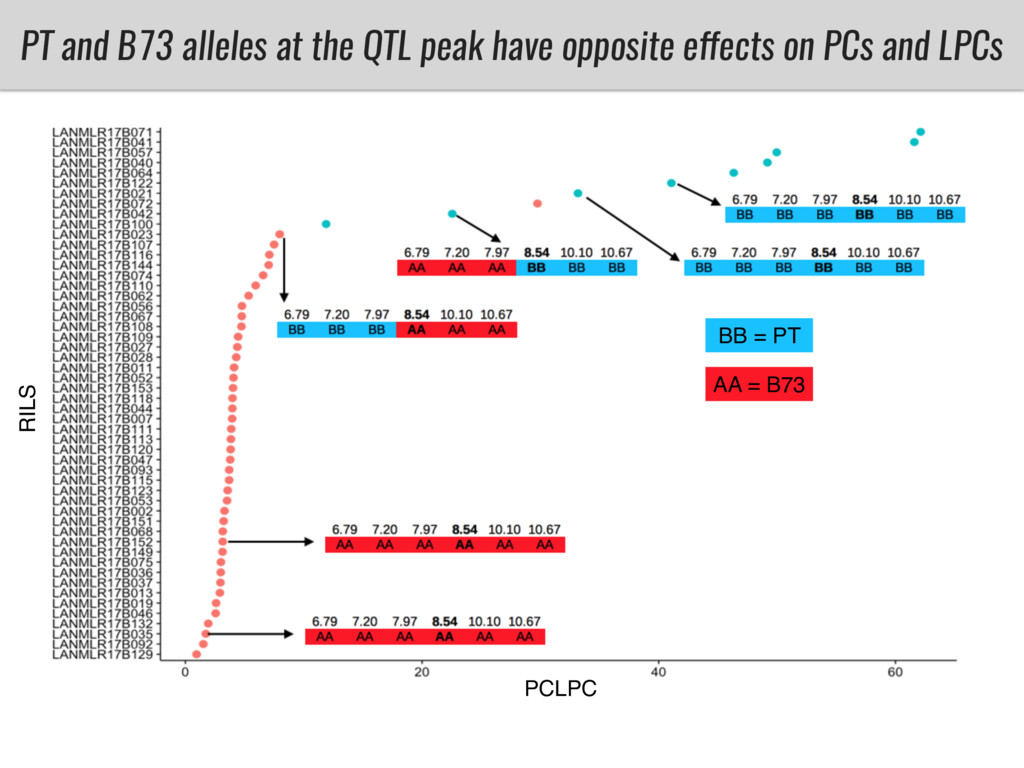
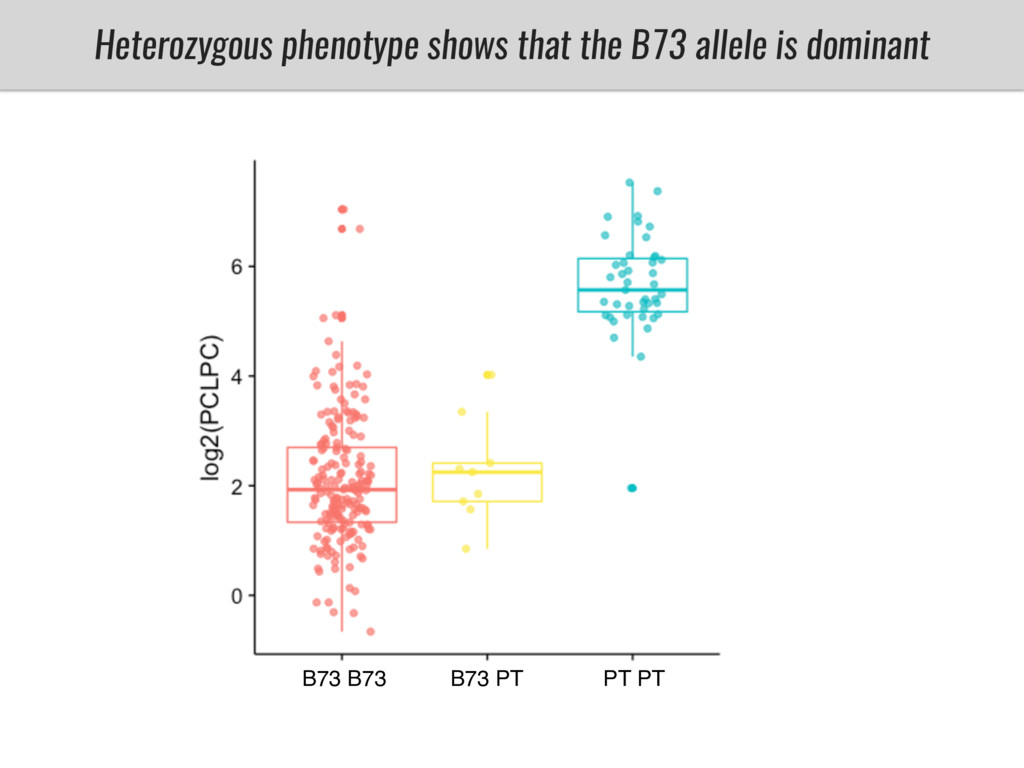
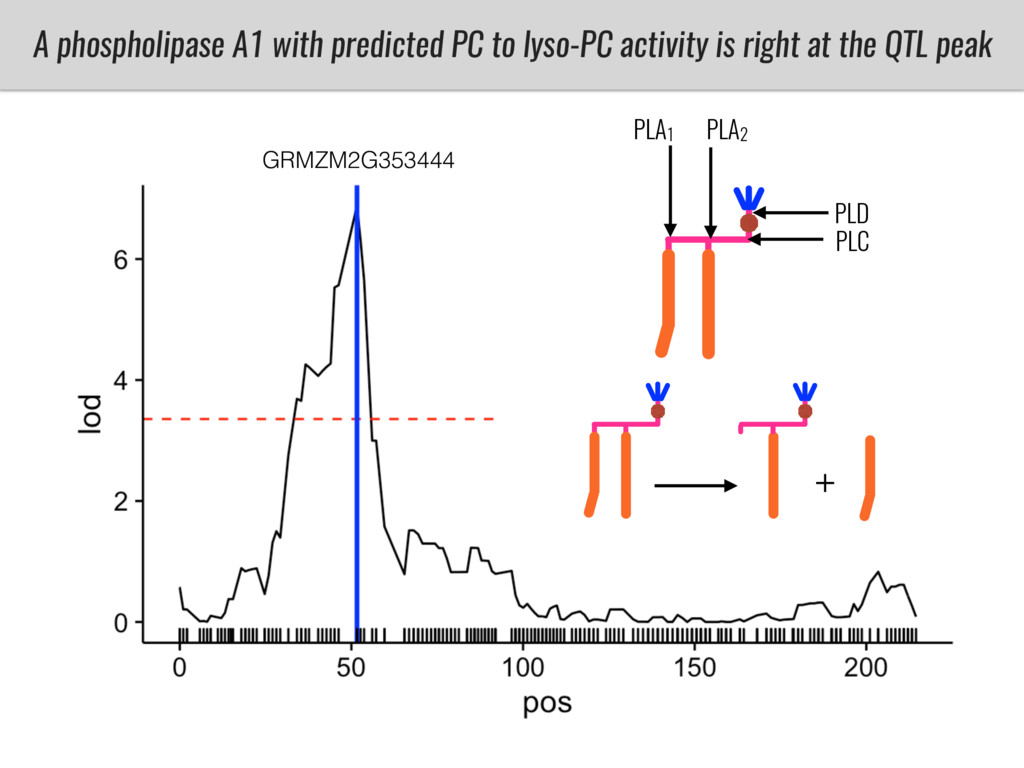
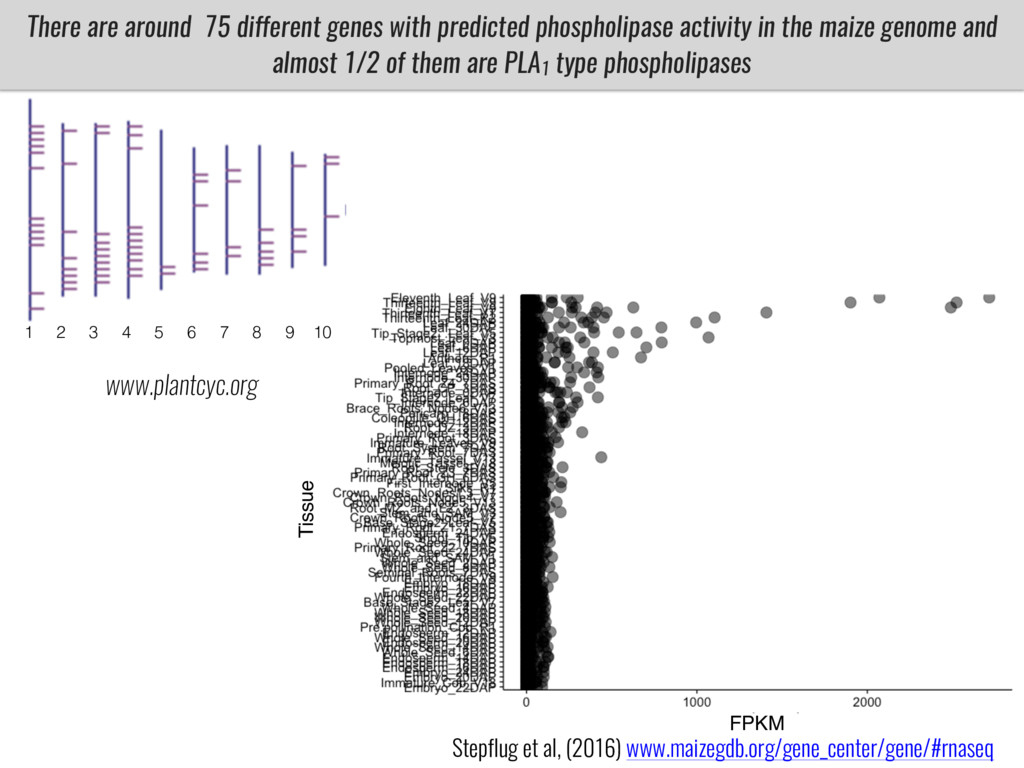
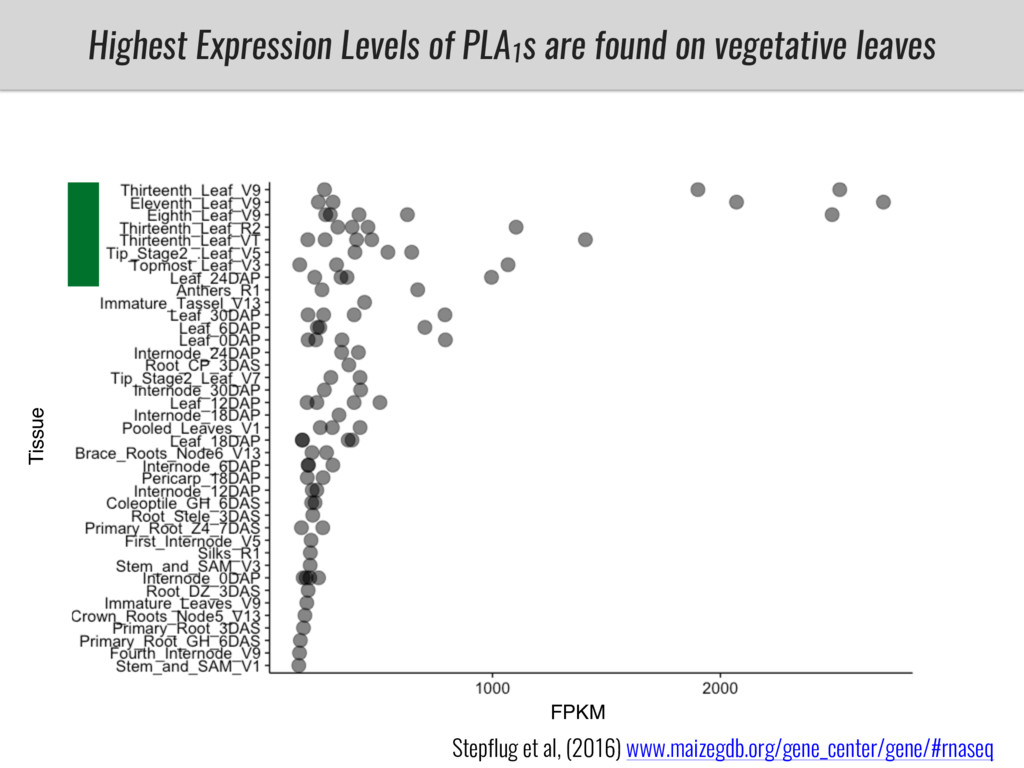
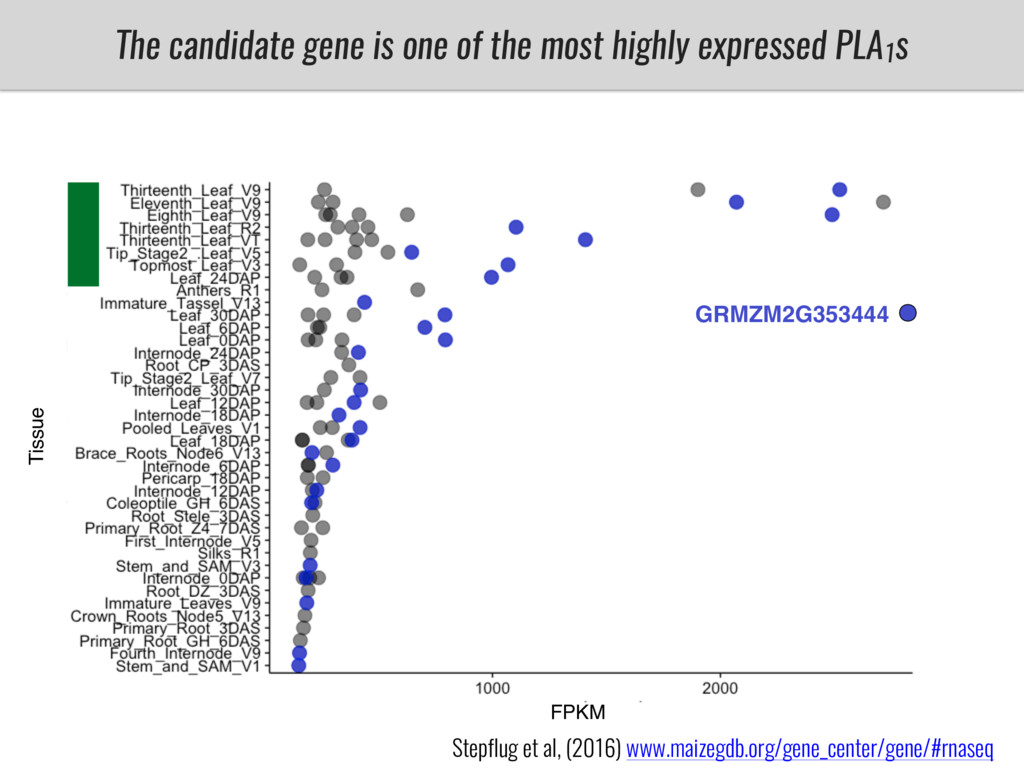
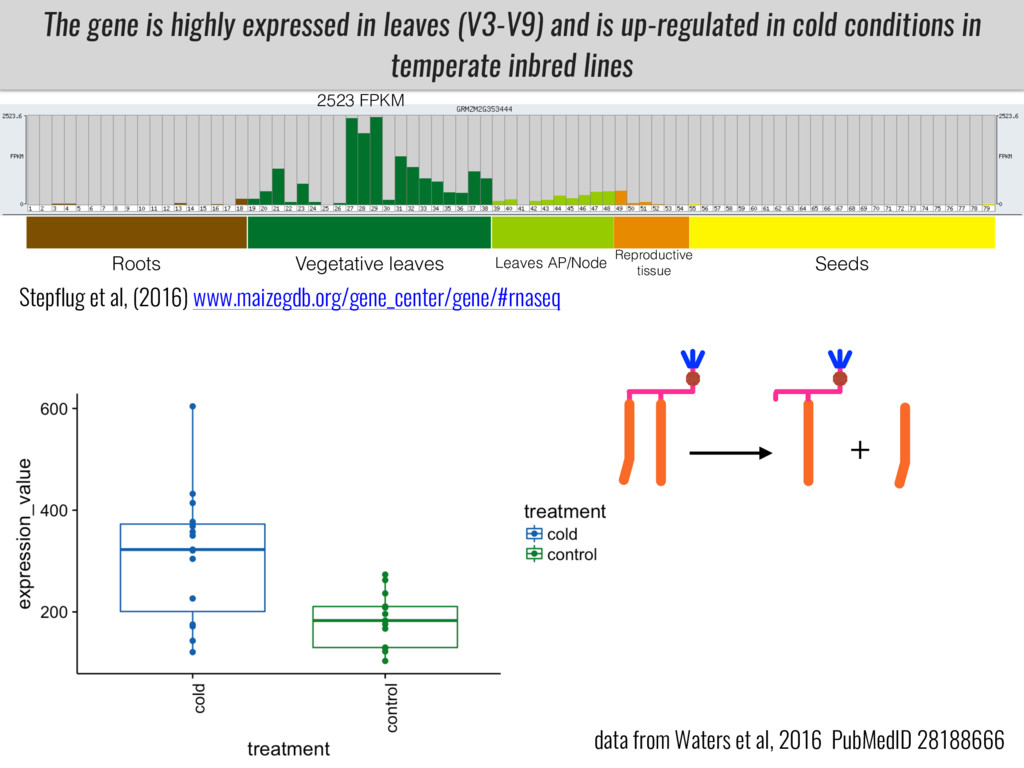
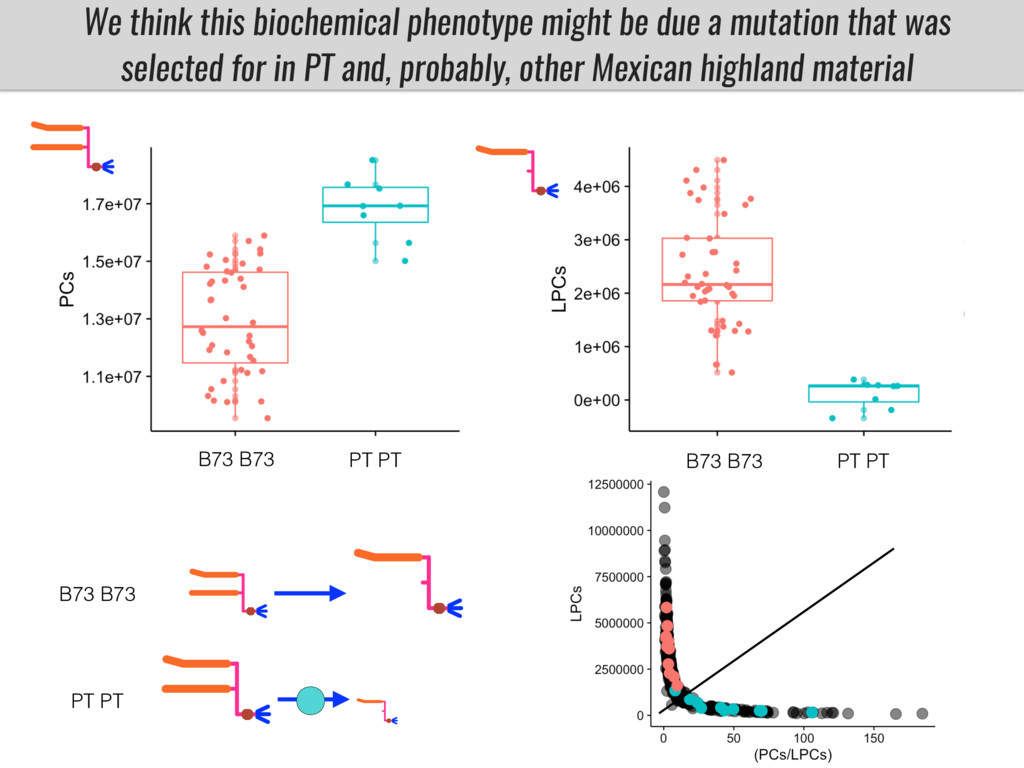
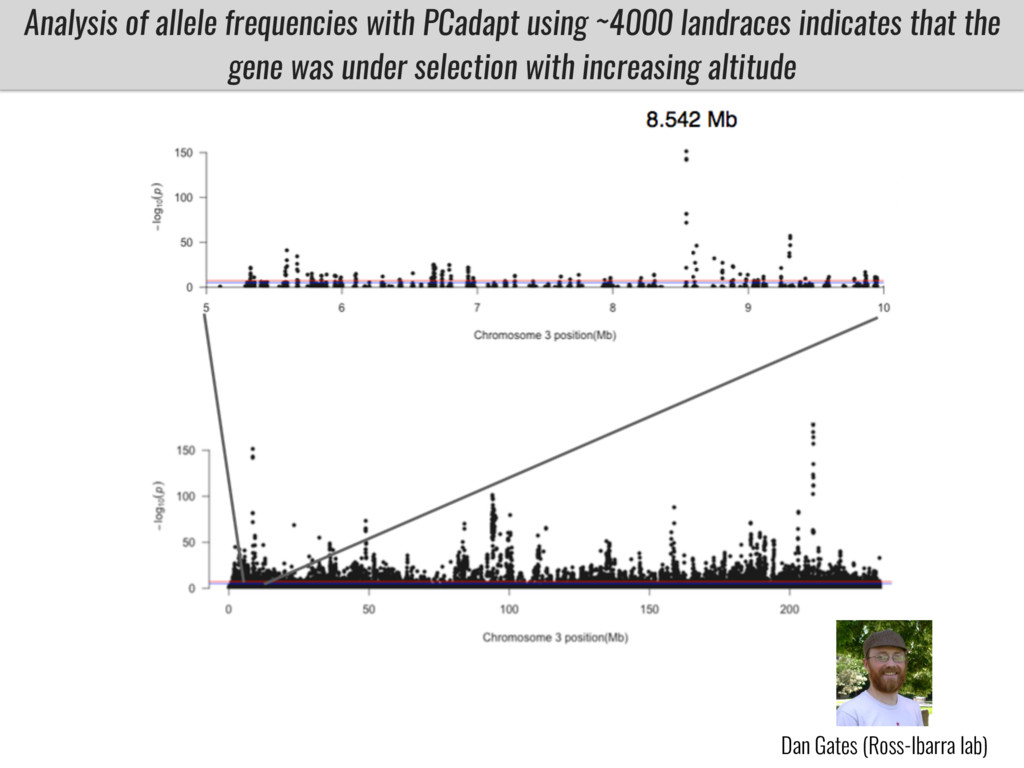
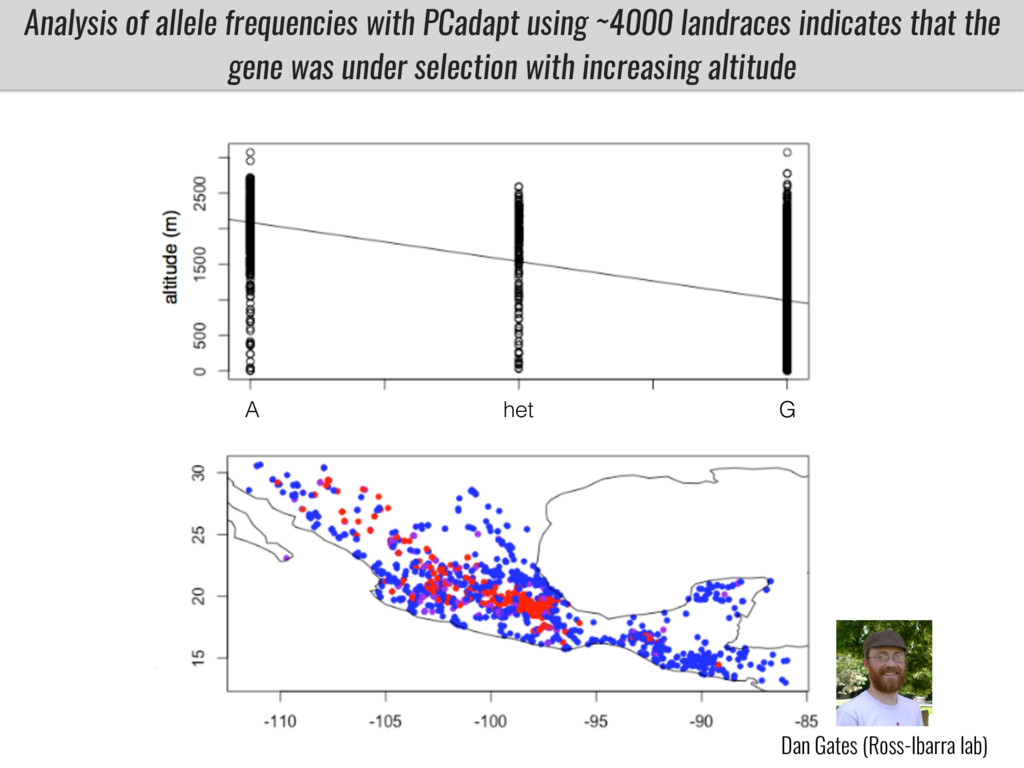
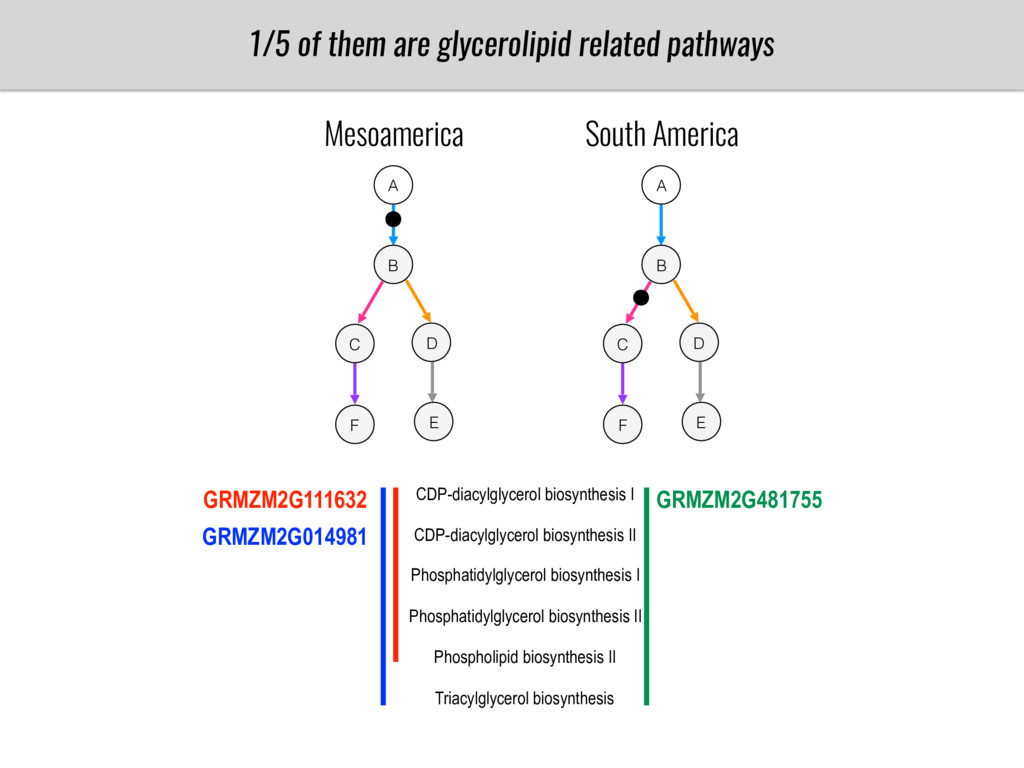
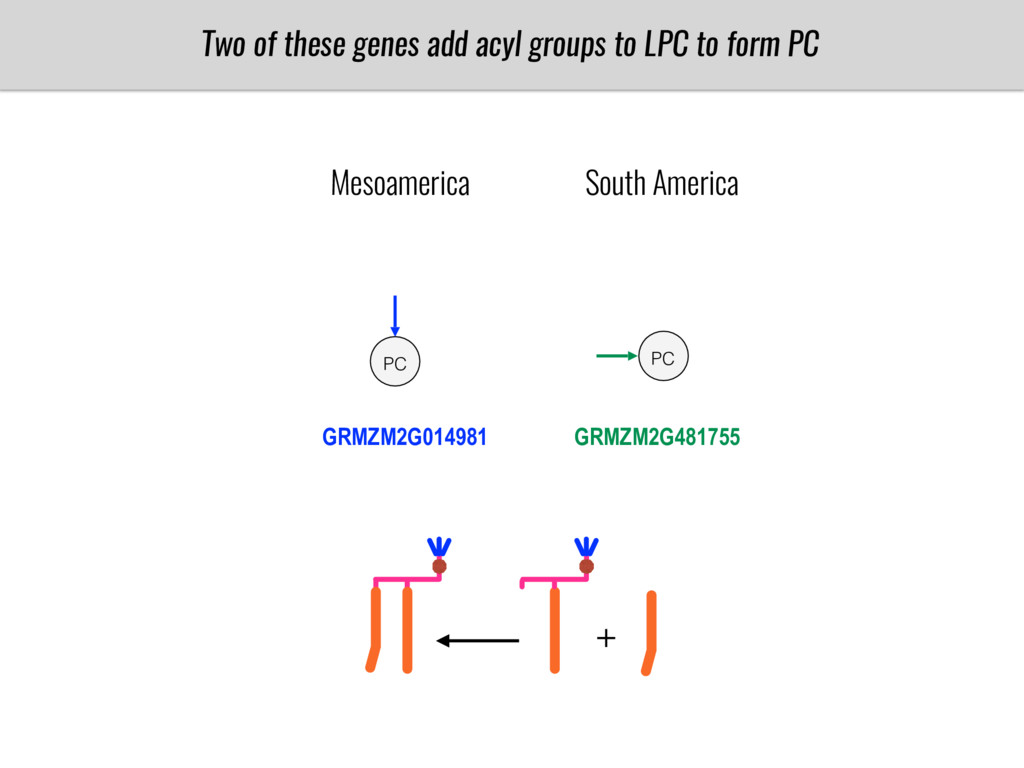
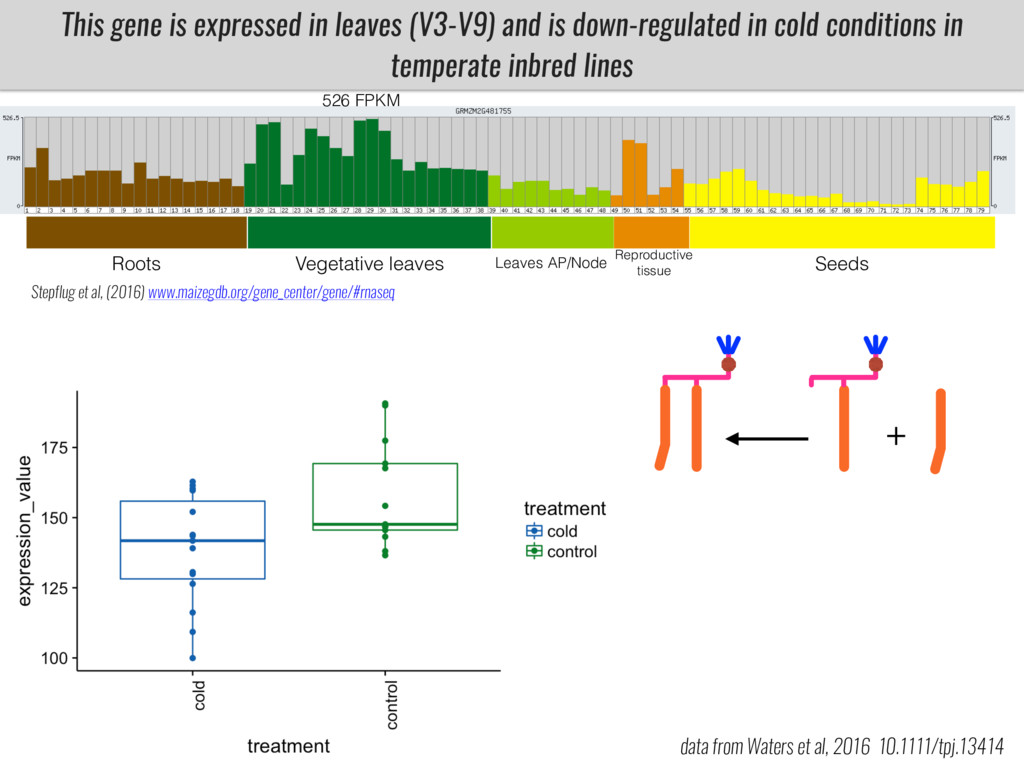
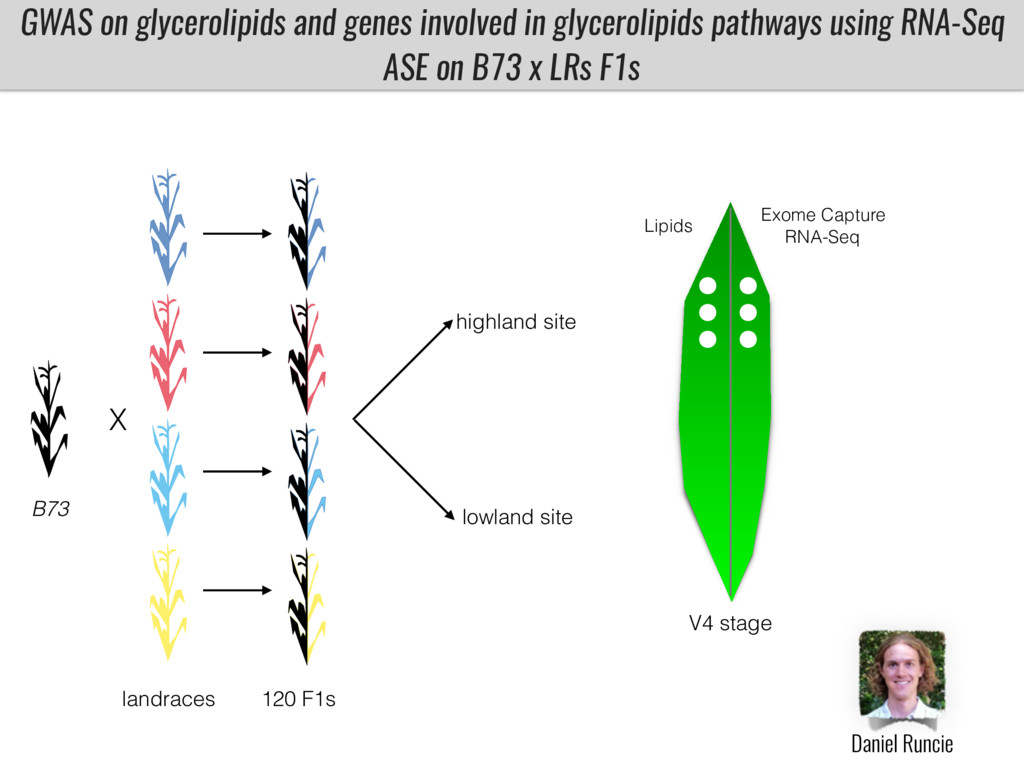
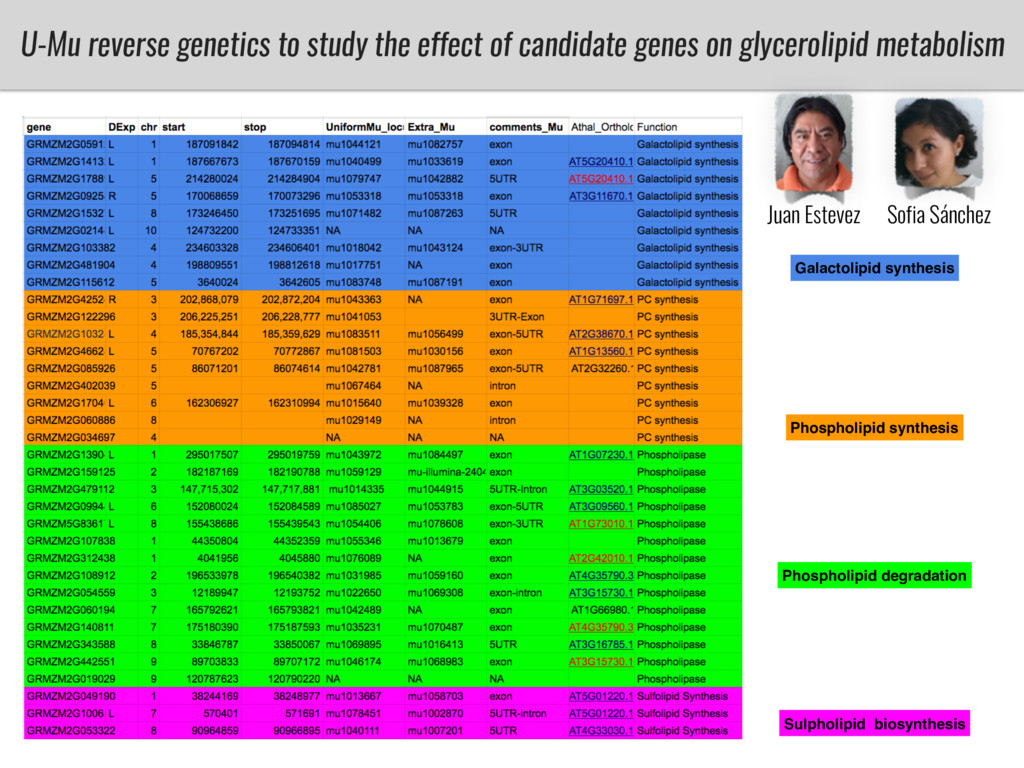
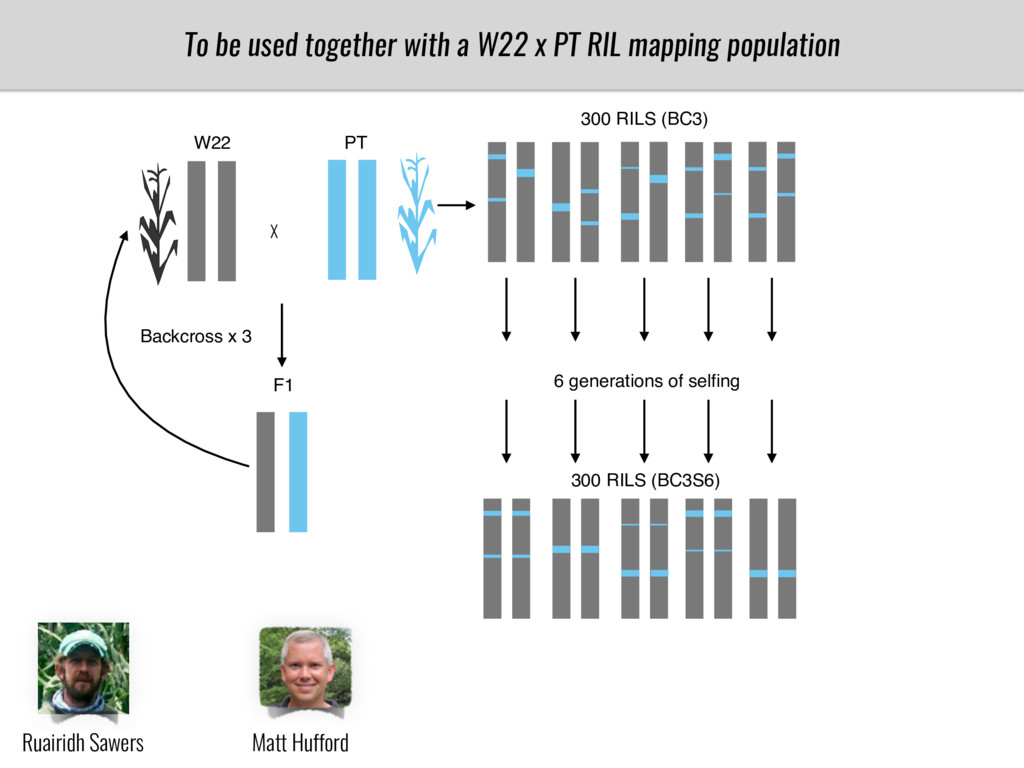
I will present data on a couple of common garden experiments in highland and lowland Mexican field sites where we grew a maize RIL mapping population and a 120 landrace diversity panel composed of highland and lowland maize. From these plants we were able to identify around 125 glycerolipid species that we are using in different ways to to identify loci involved in glycerolipid metabolism I will present these results and in particular on an interesting QTL in ch3 where a candidate phospholipase gene seems to be responsible of phosphatidylcholine (PC) to lysophosphatidylcholine (lyso-PC) conversion.
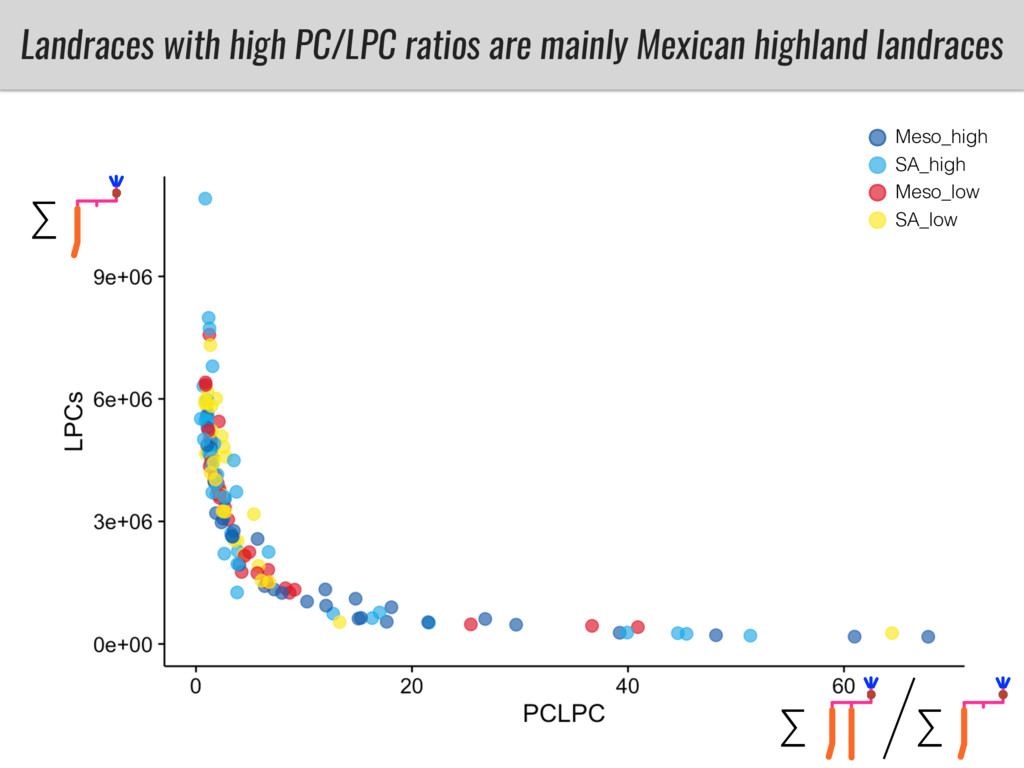
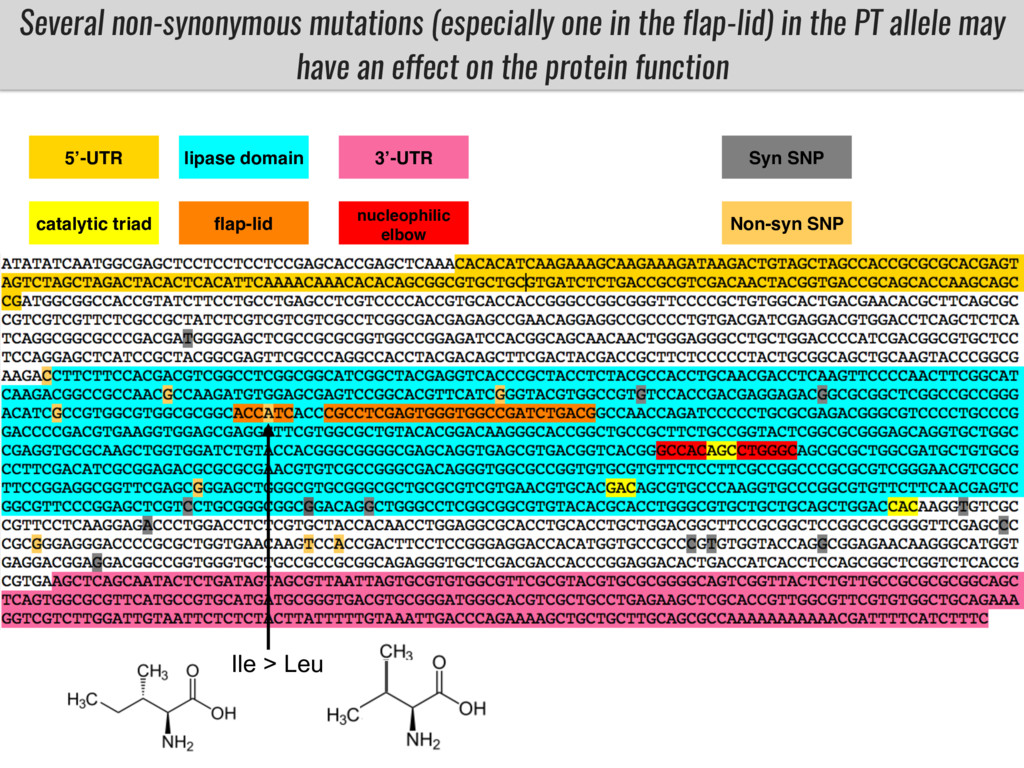
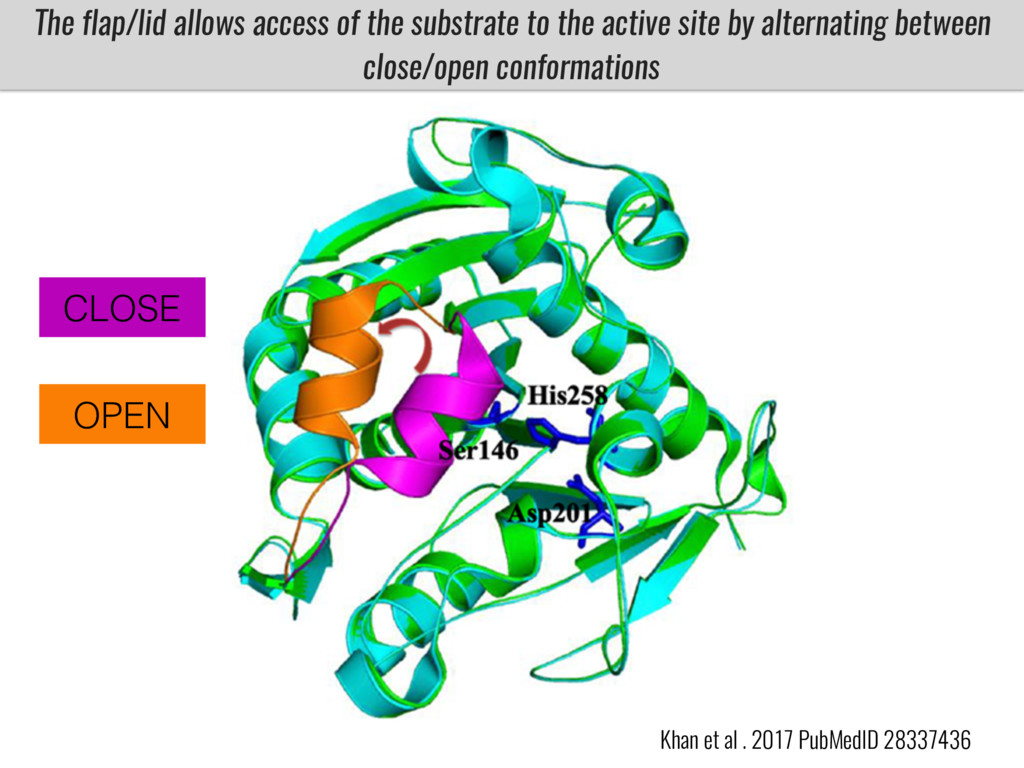
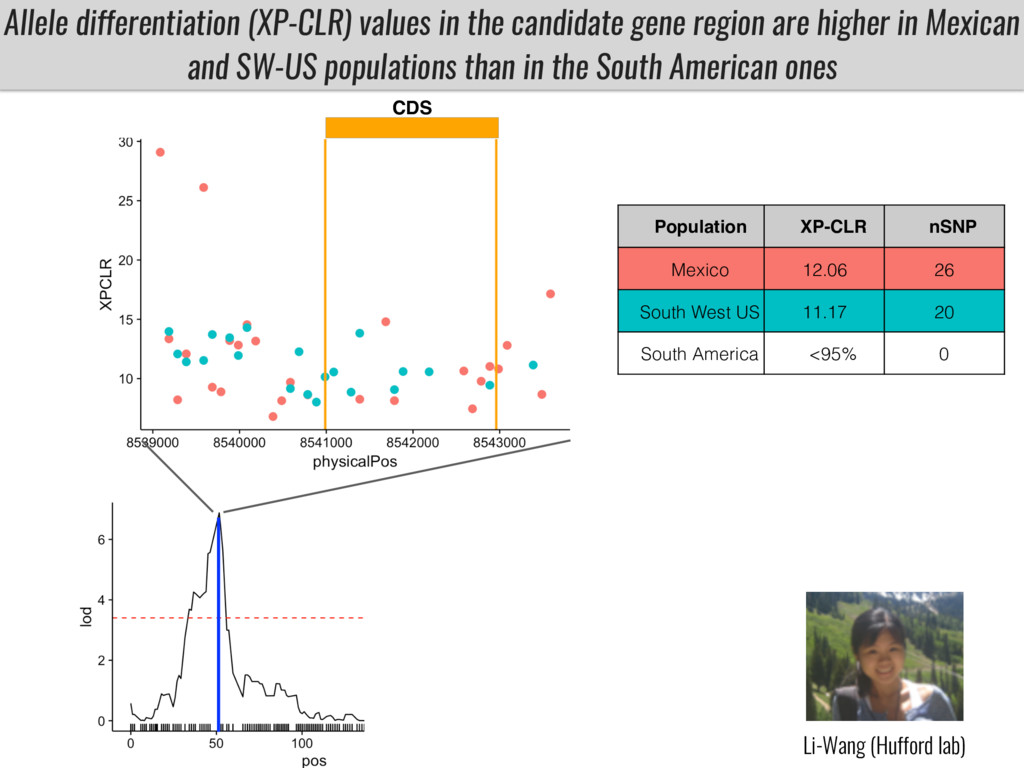
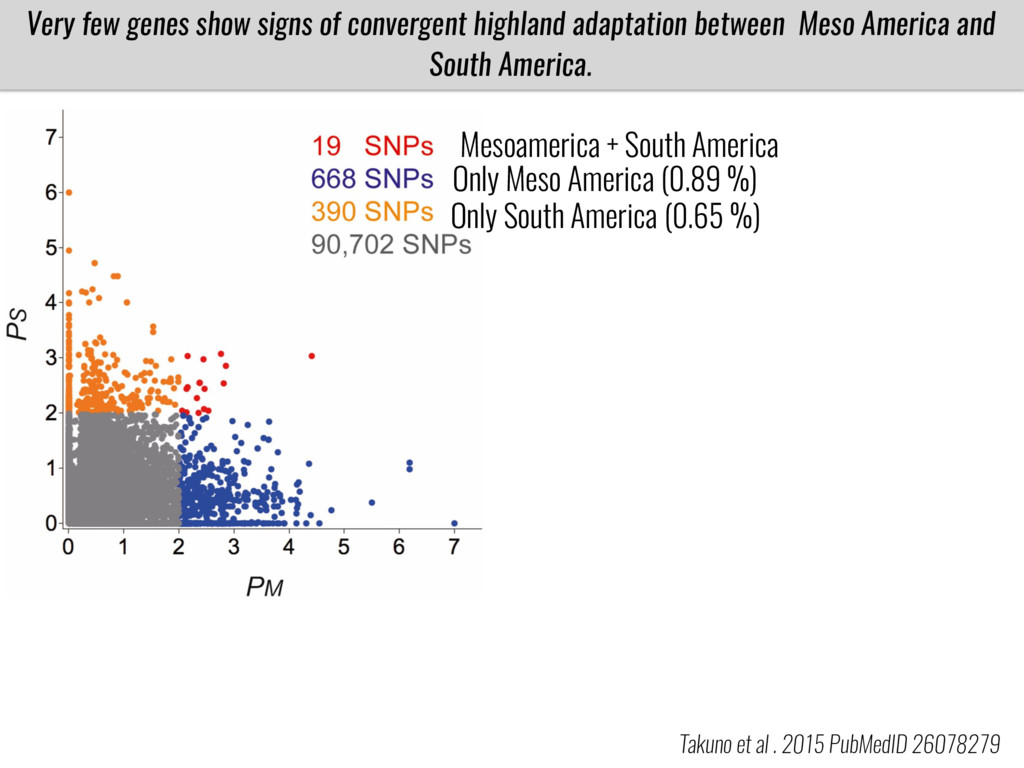
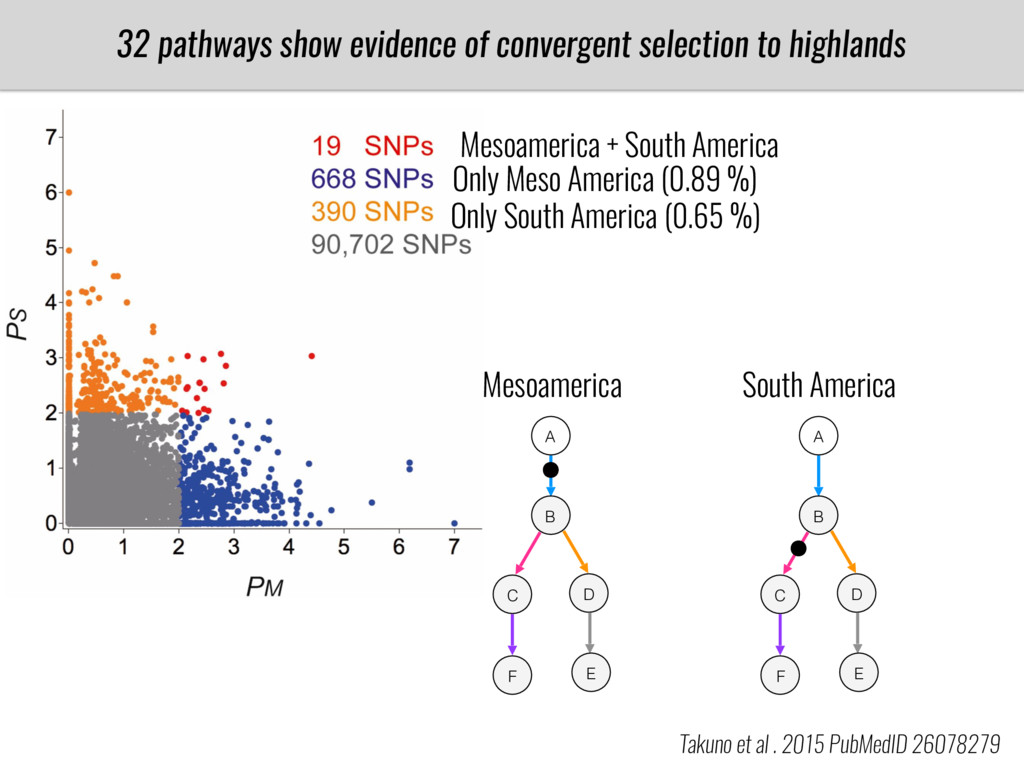
Our phenotypic data together with population differentiation data suggests that genes controlling PC to lyso-PC conversion were under selection during the process of maize adaptation to the highlands. We are now trying to understand to what extent balancing PC to lyso-PC species might be important to local adaptation in the highlands, the possible convergence of this phenotype in Mexican and South American highland maize and the possibility to apply this knowledge understand how maize adapts to different temperatures and phosphorus levels.
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
![pH ~ 5.1 [P] ~ 4 ppm 3570 masl](https://files.speakerdeck.com/presentations/bedde92950084697b8f72ee70c3b105b/slide_19.jpg){kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}