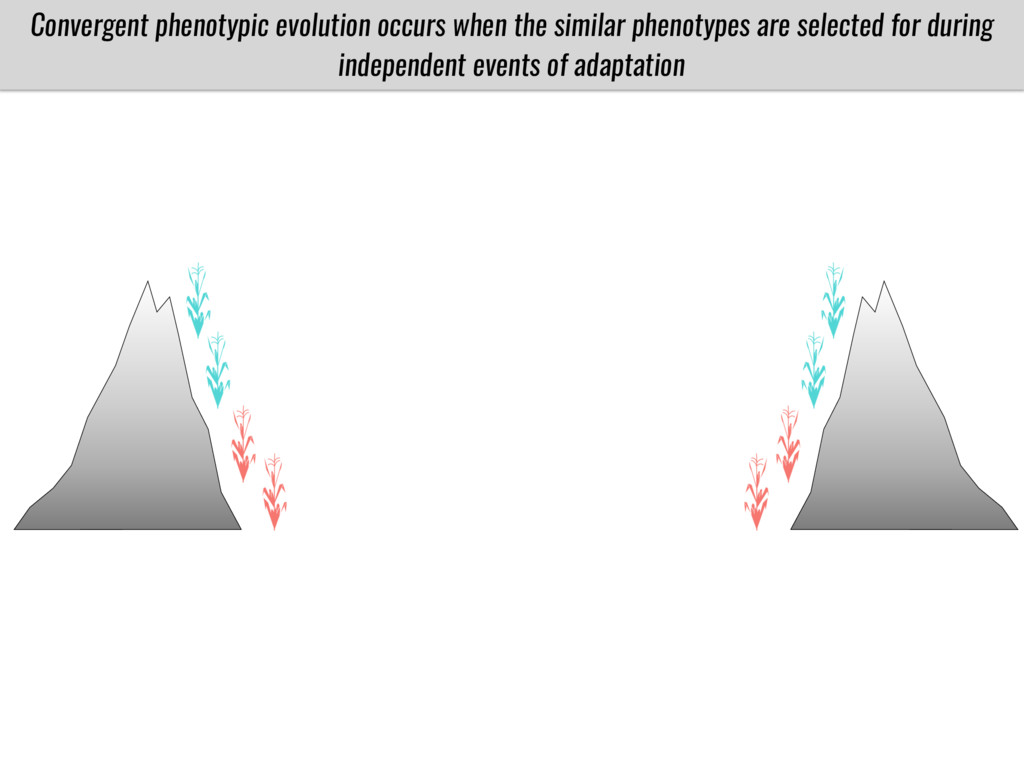
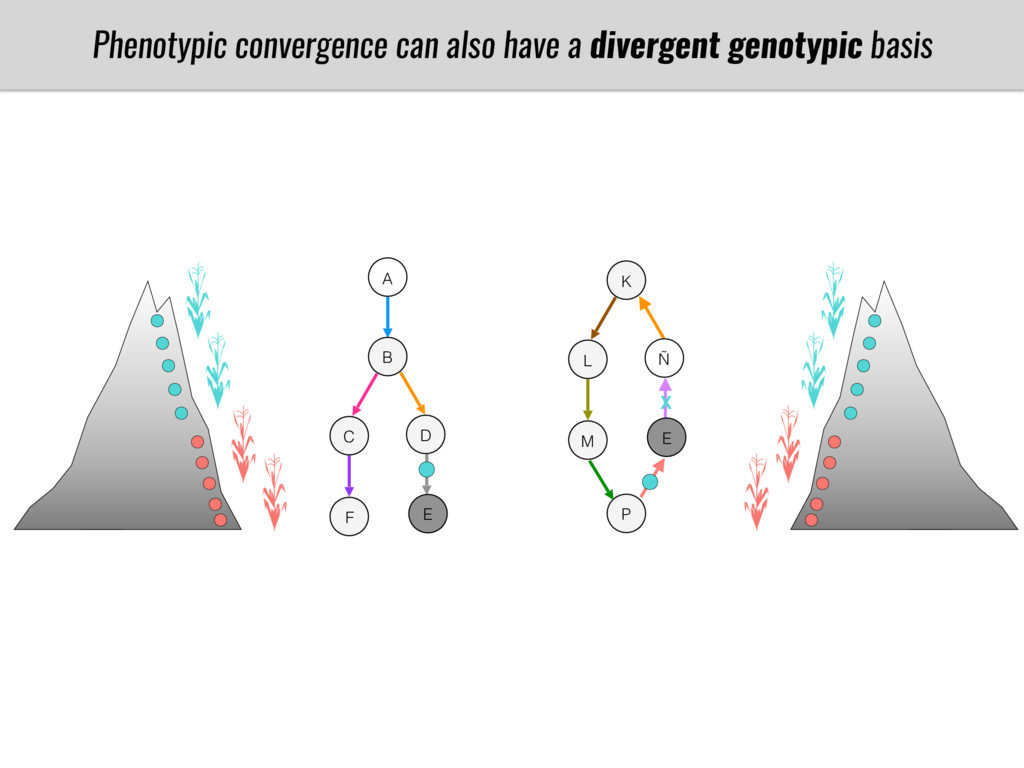
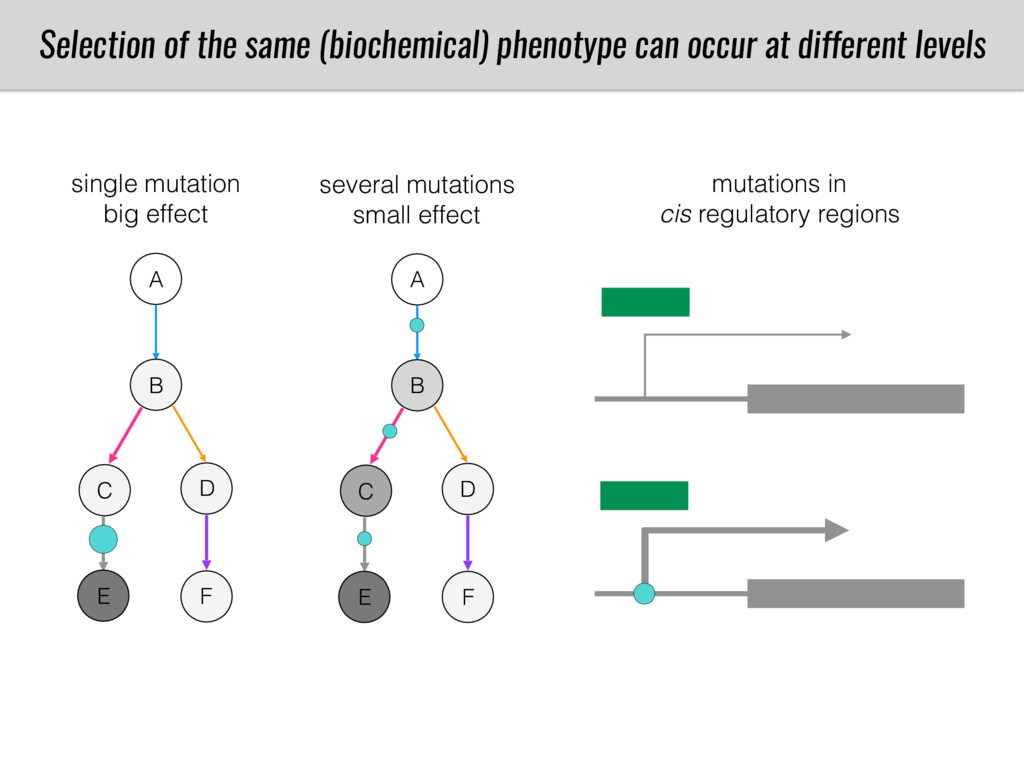
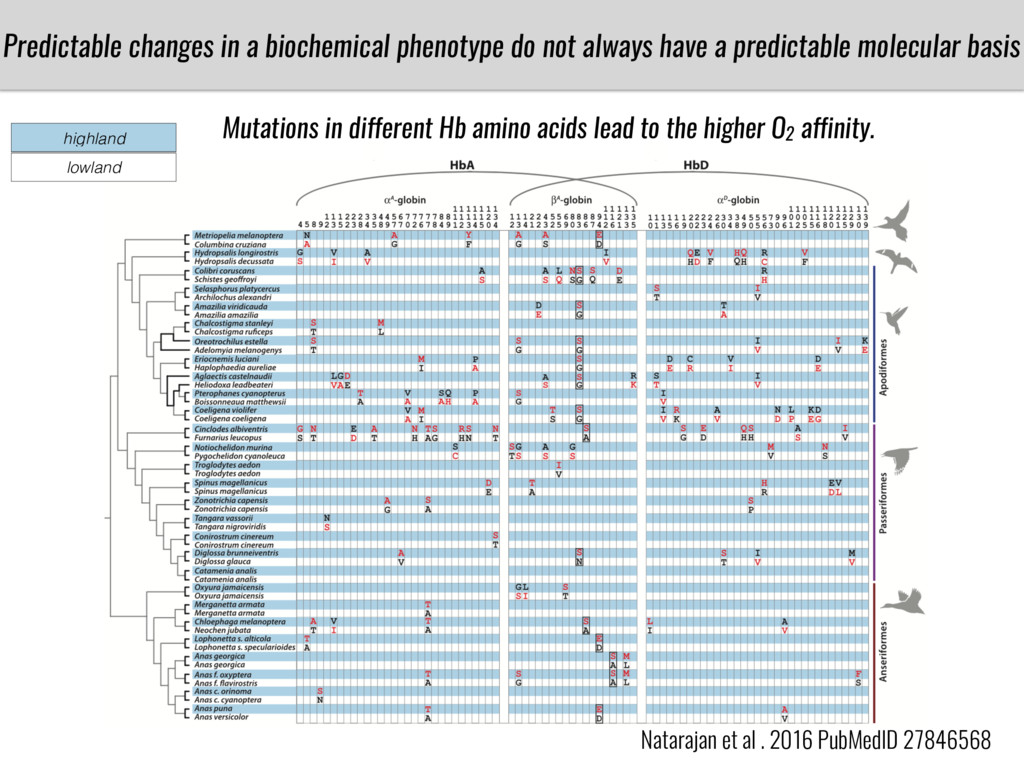
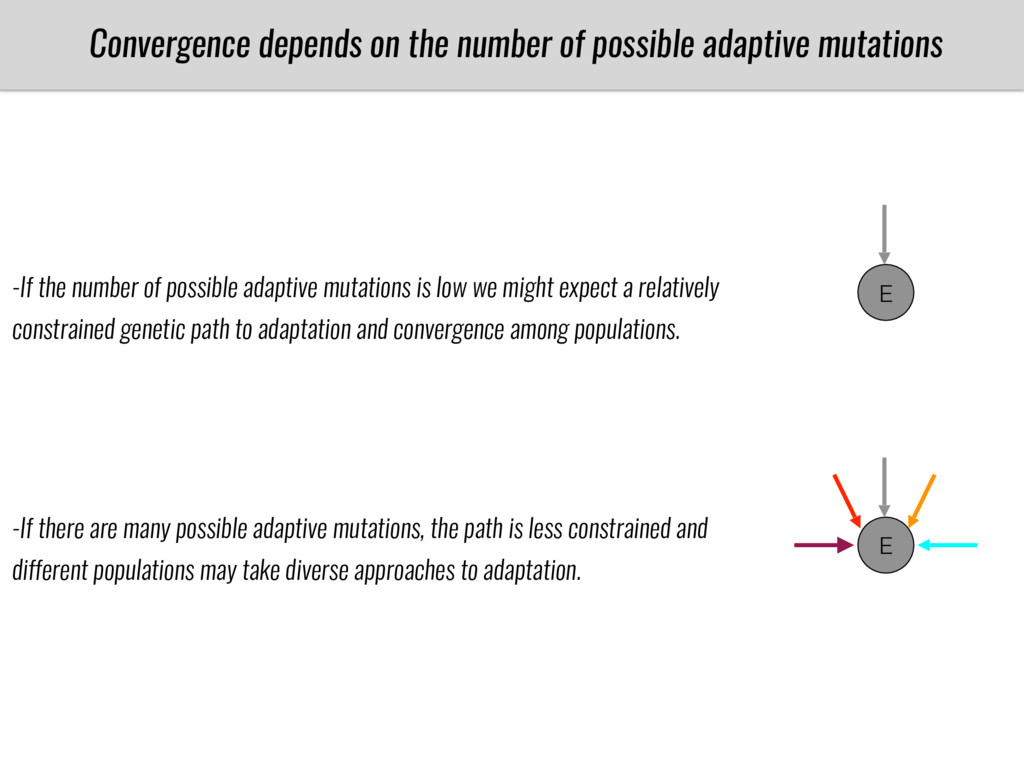
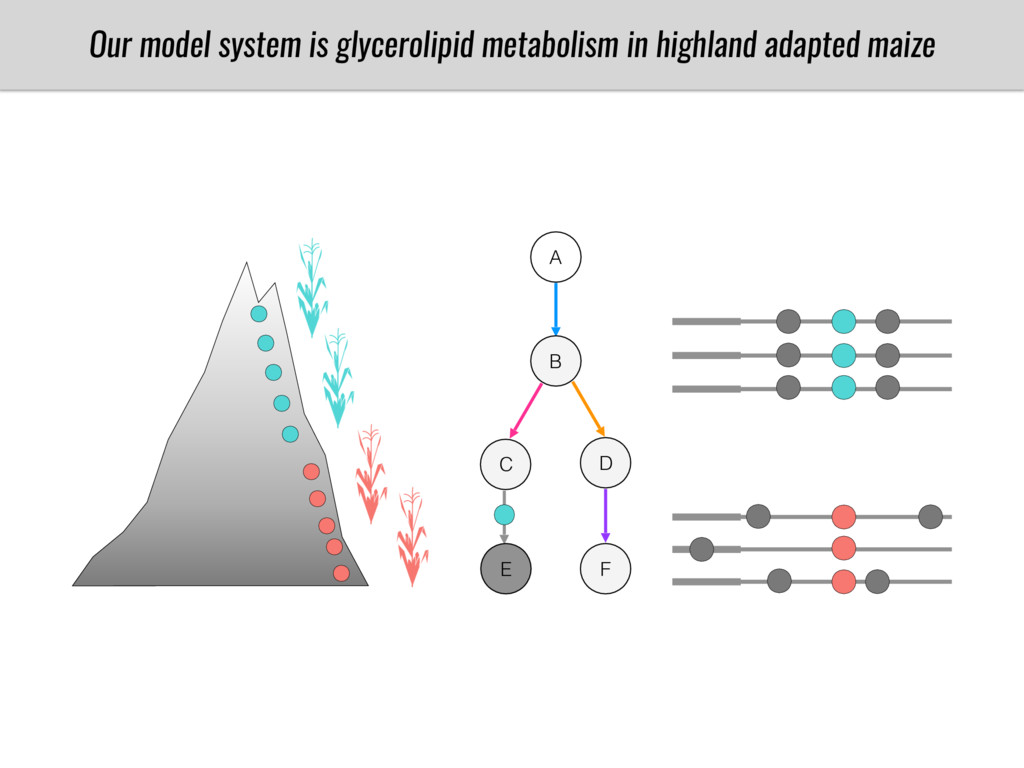
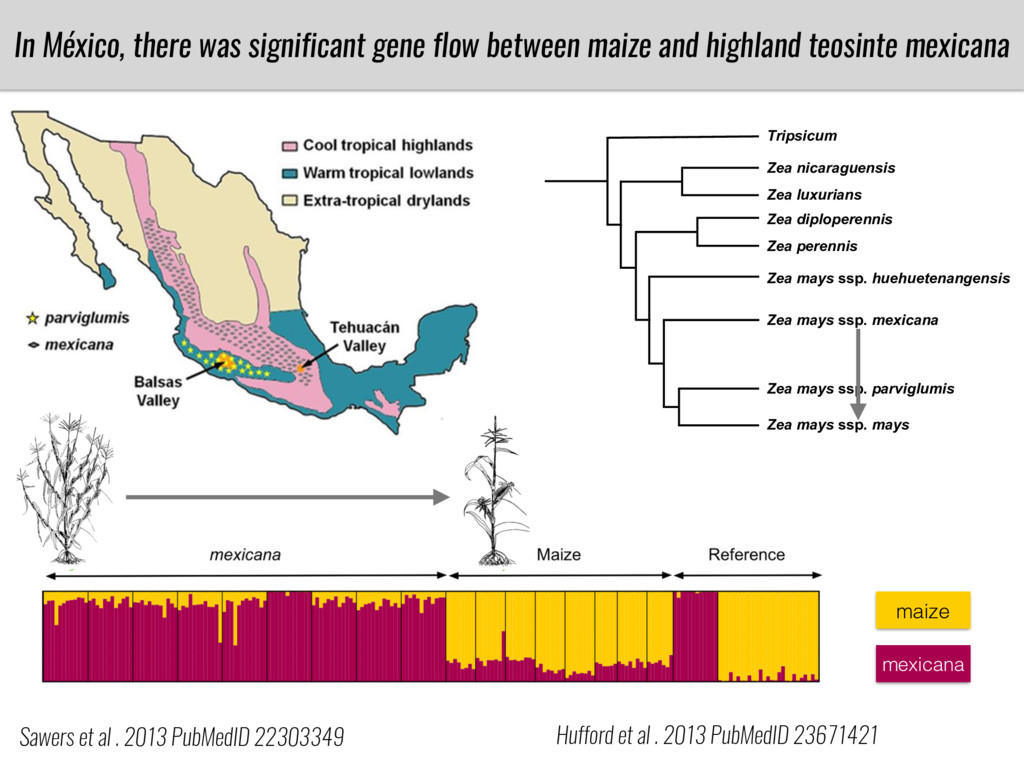
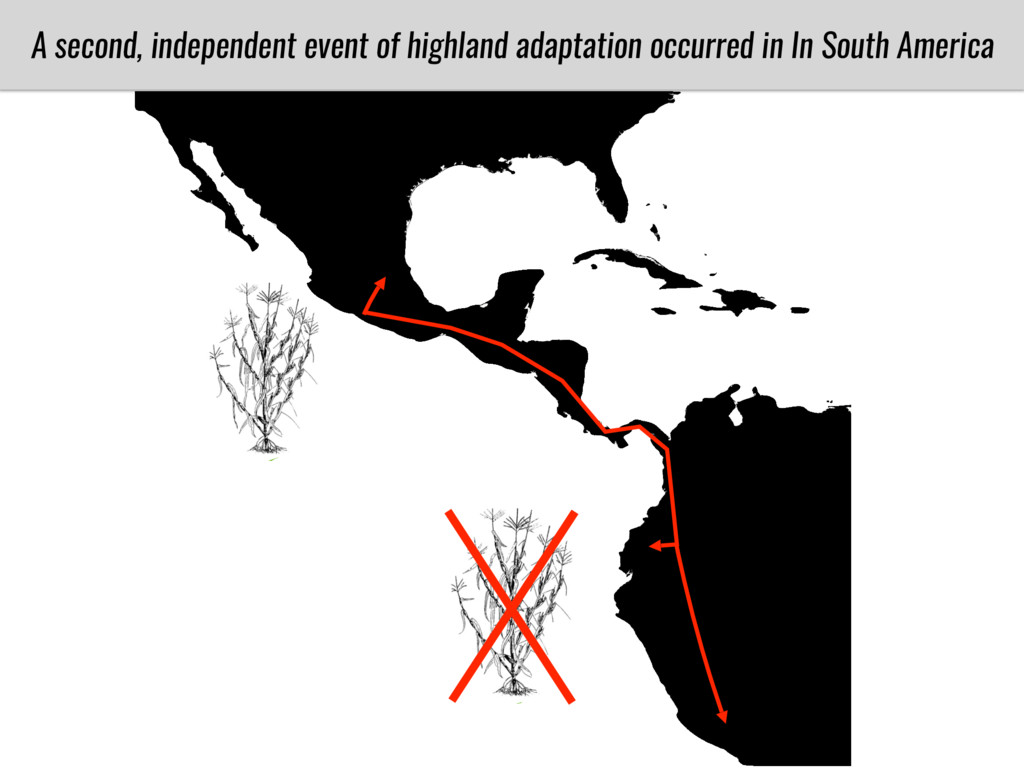
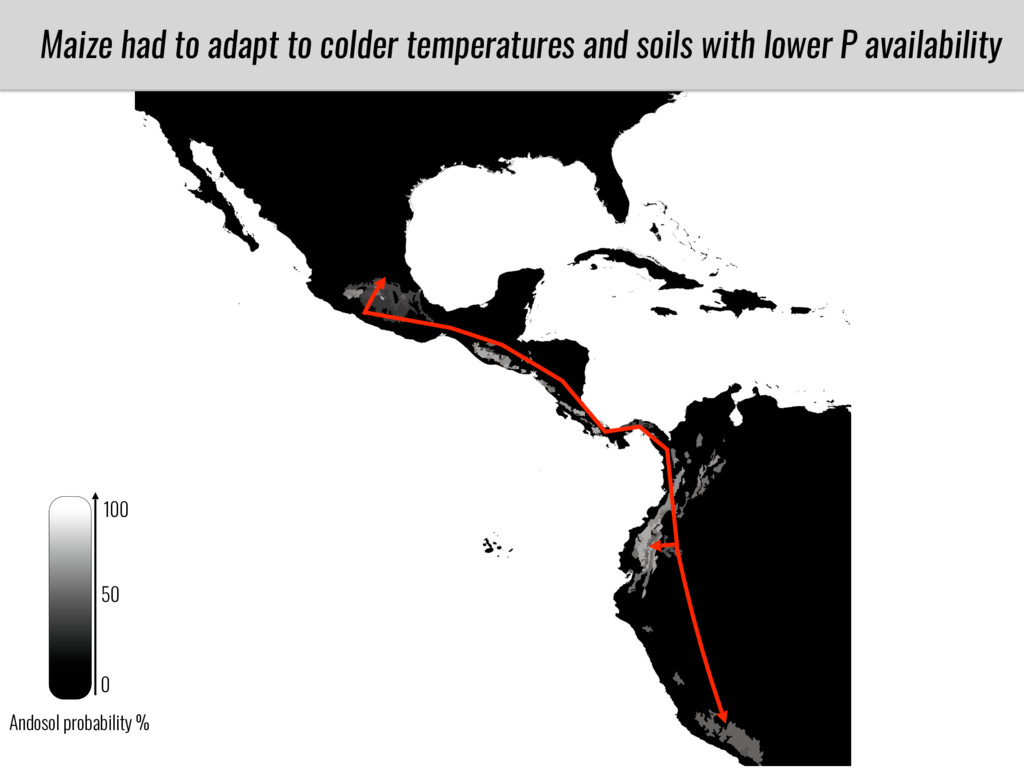
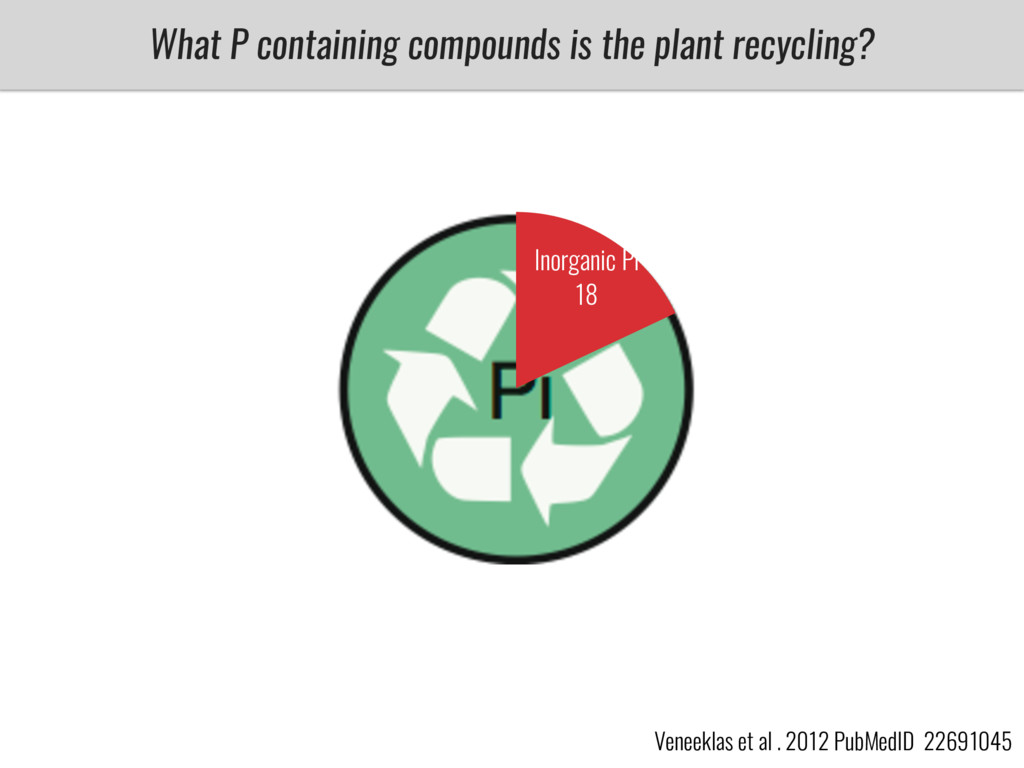
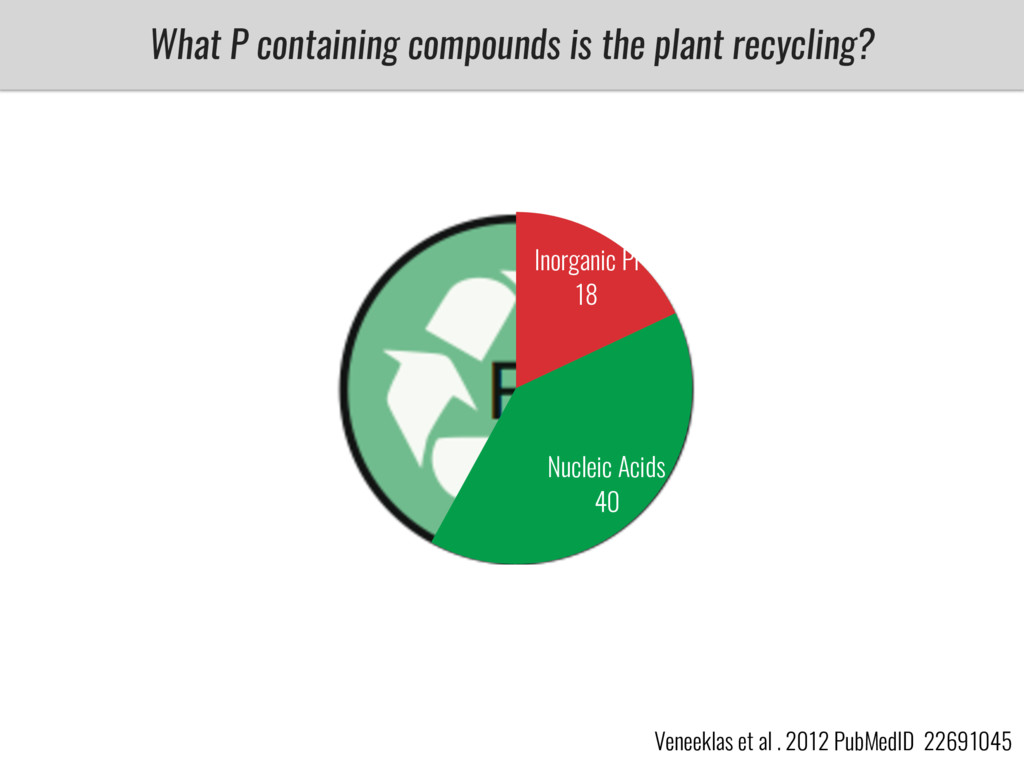
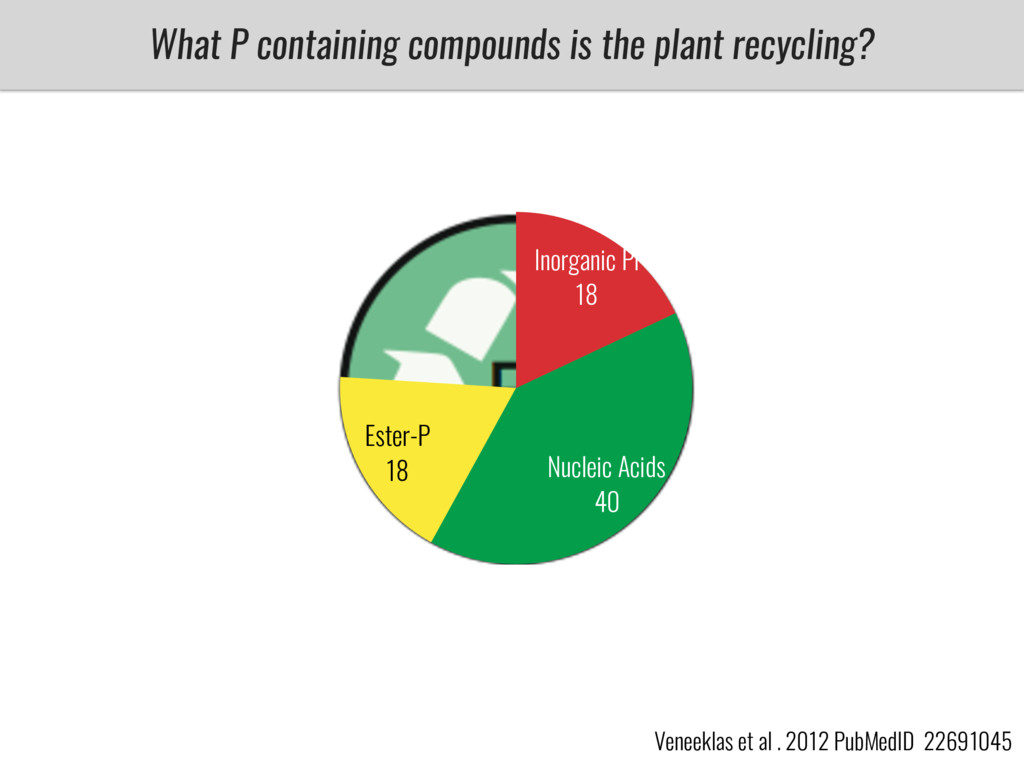
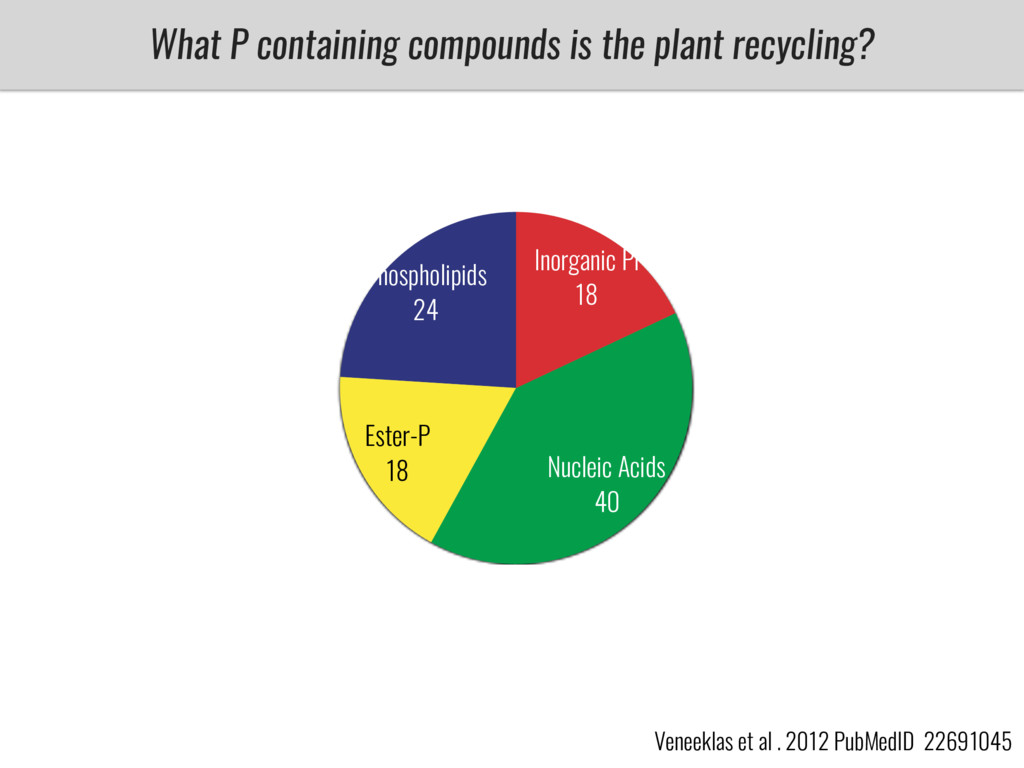
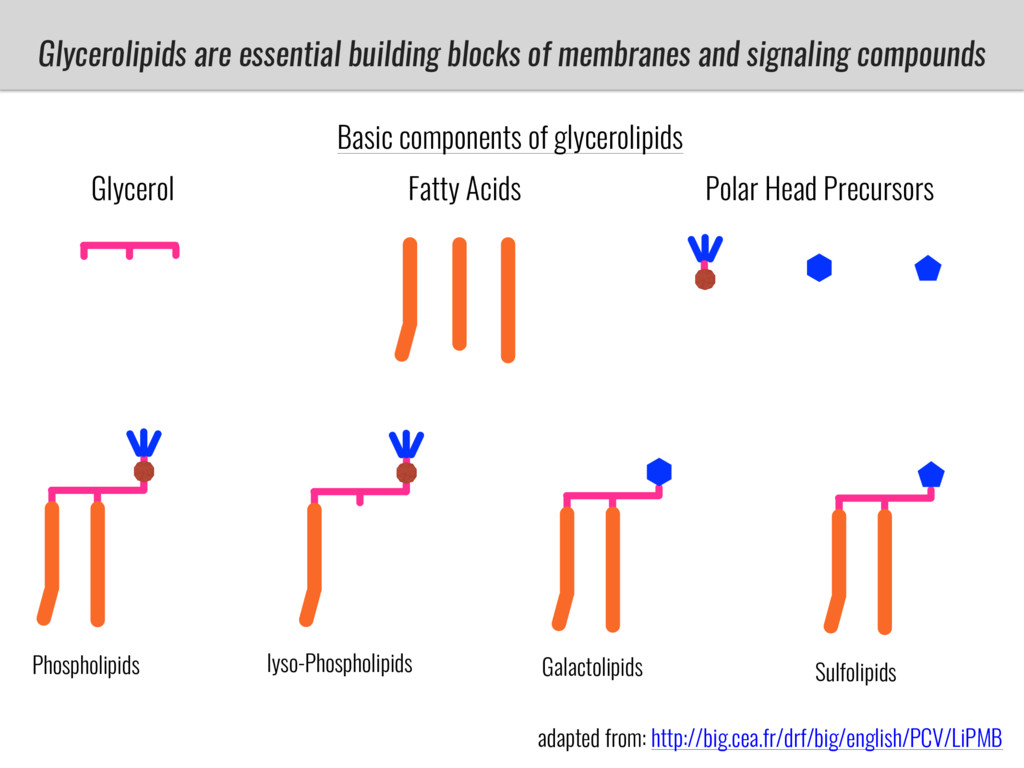
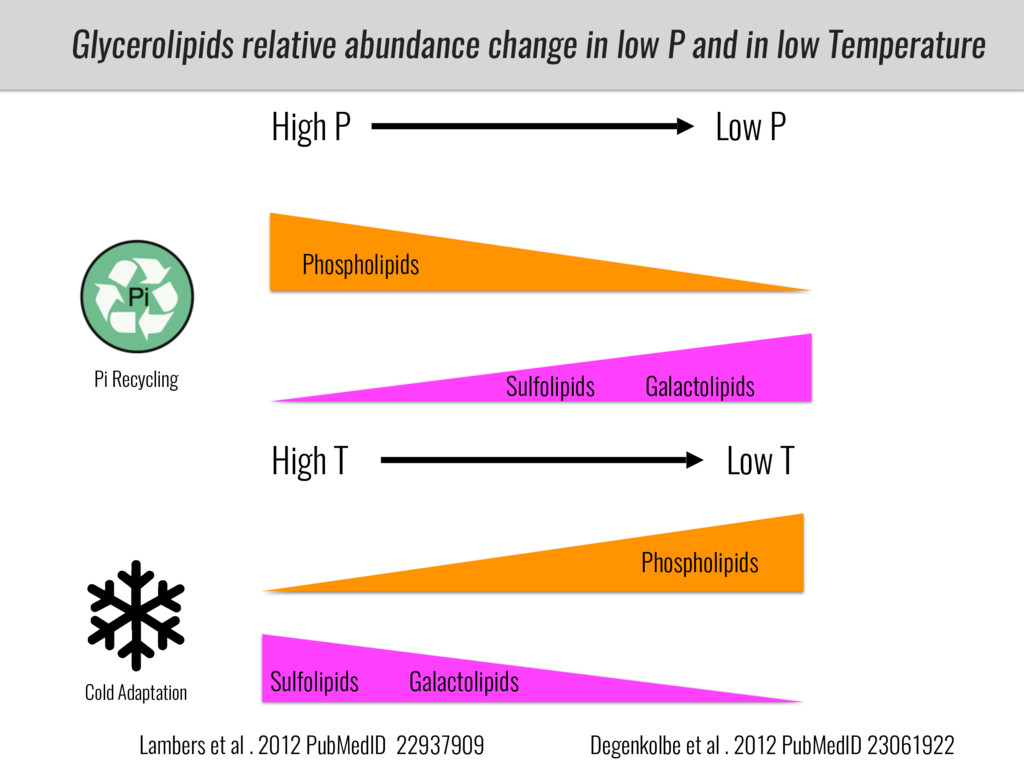
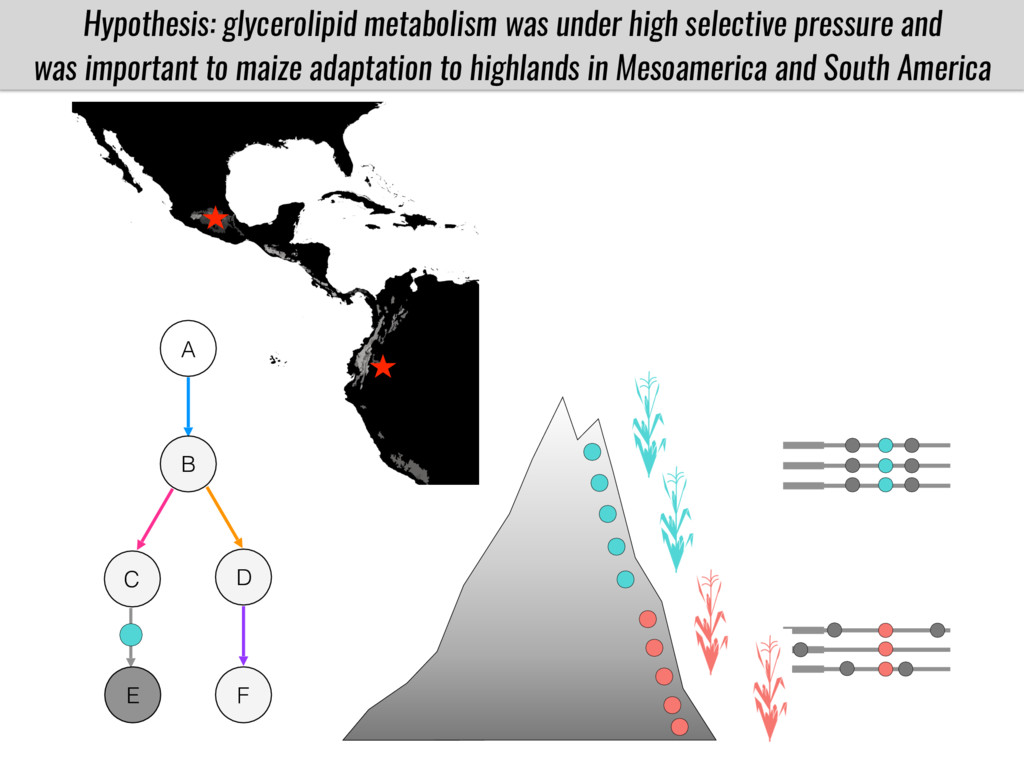
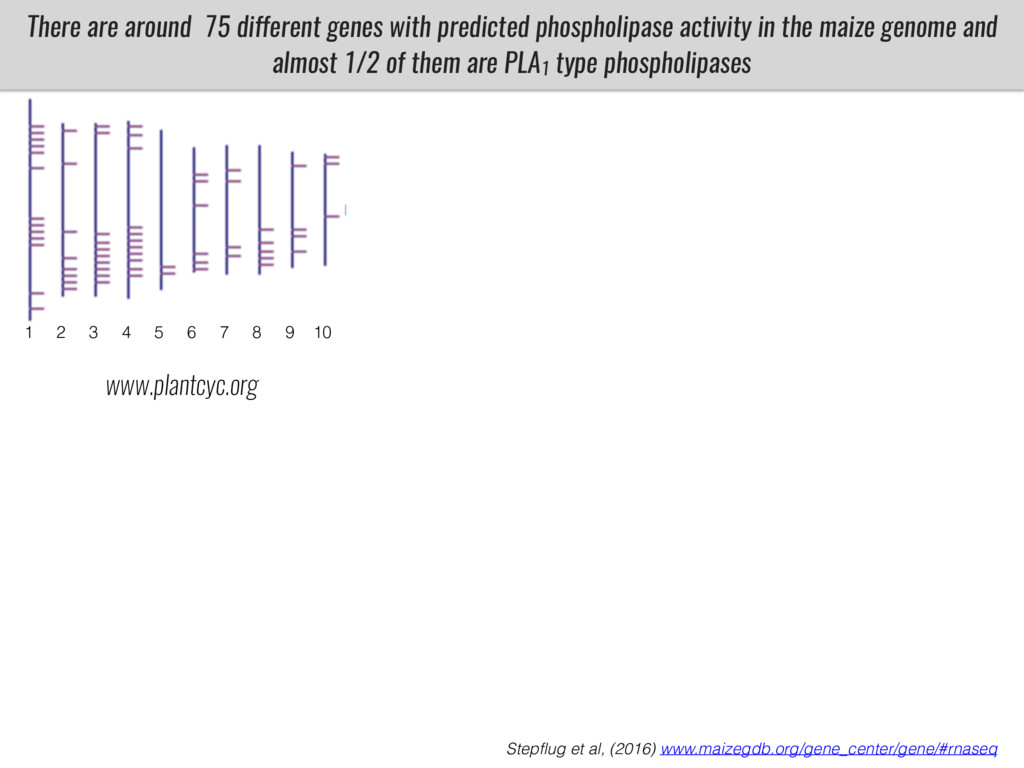
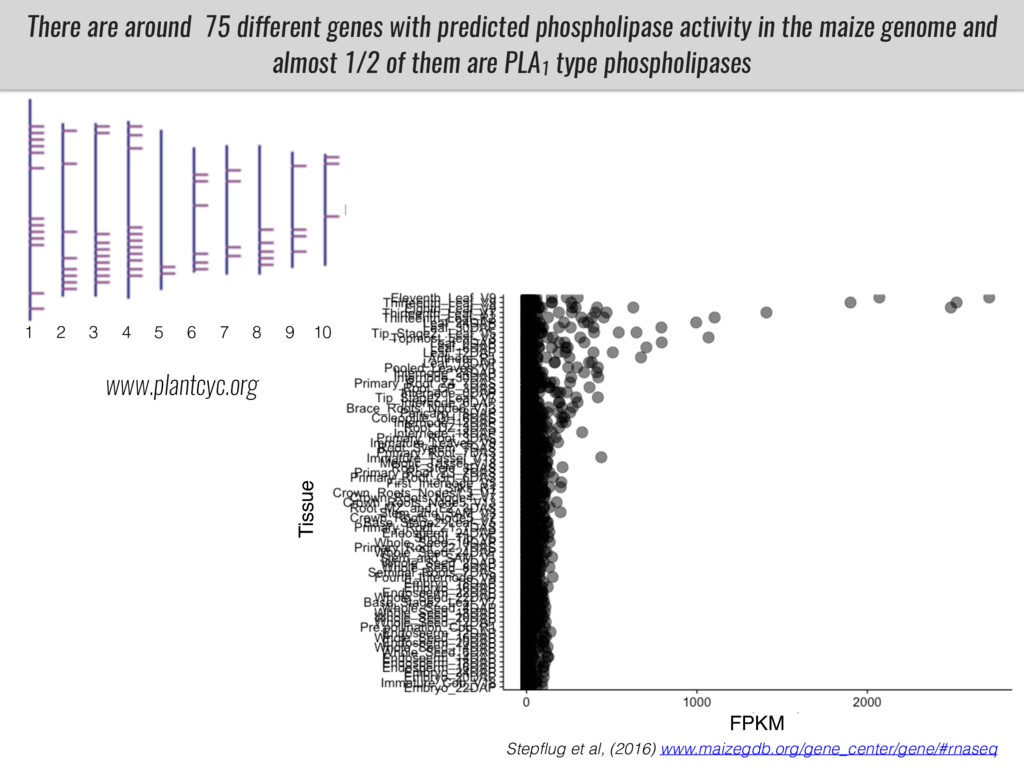
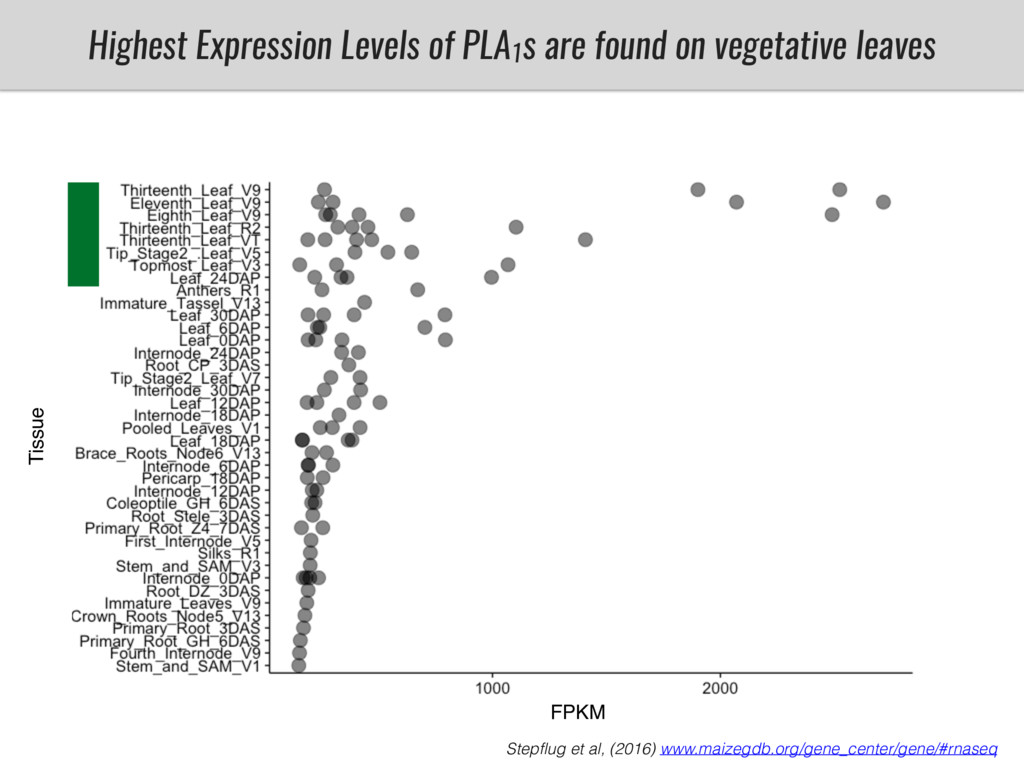
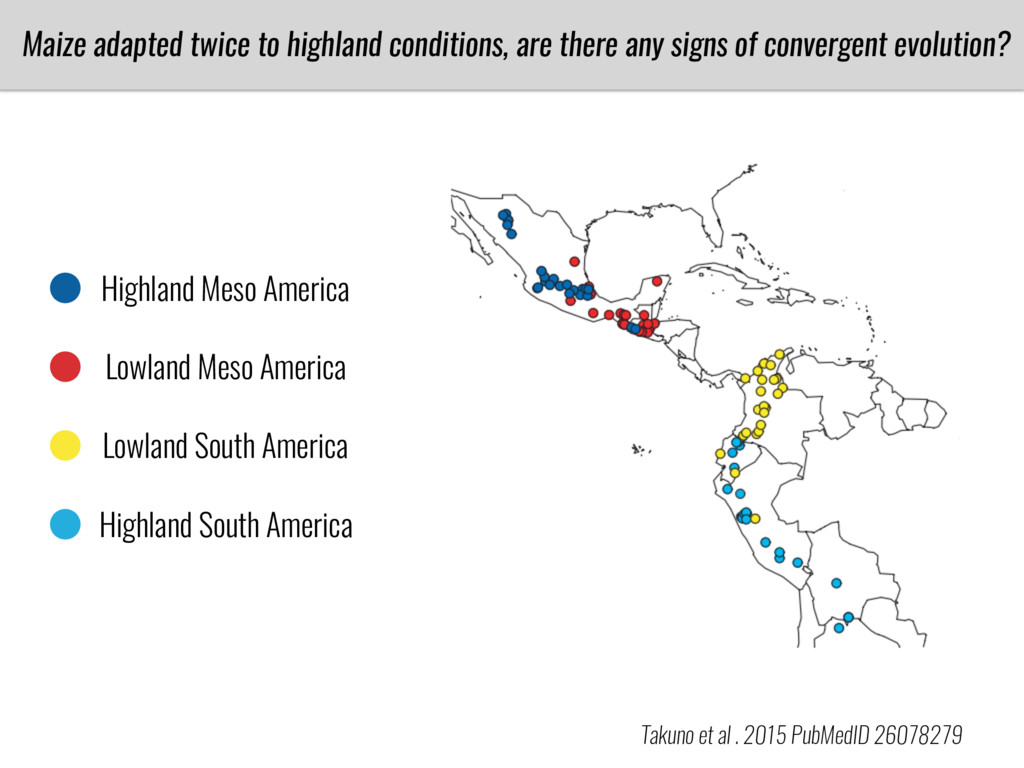
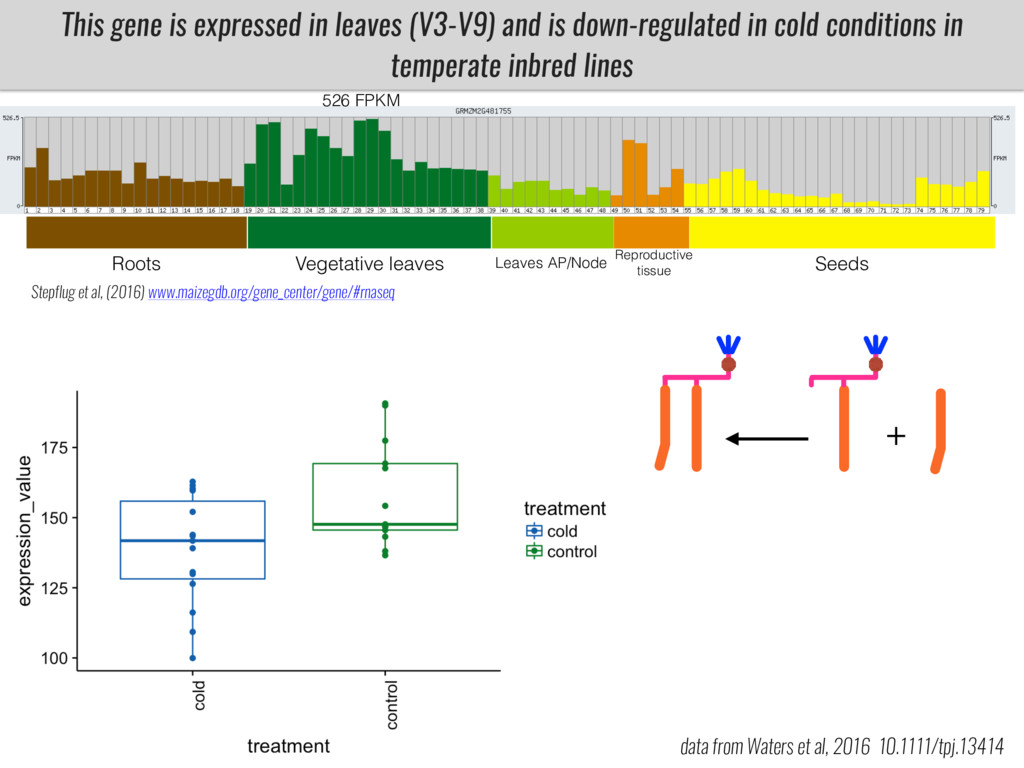
After domestication from lowland teosinte in Mexican subtropic maize colonized the highlands of both Mexico and South America characterized by lower temperatures and phosphorus bioavailability. We hypothesize that low temperature and phosphorus bioavailability were major selective forces during maize adaptation to the highlands and shaped glycerolipid metabolism since low temperature and P deficiency have opposite effects on the content of glycerolipid species.
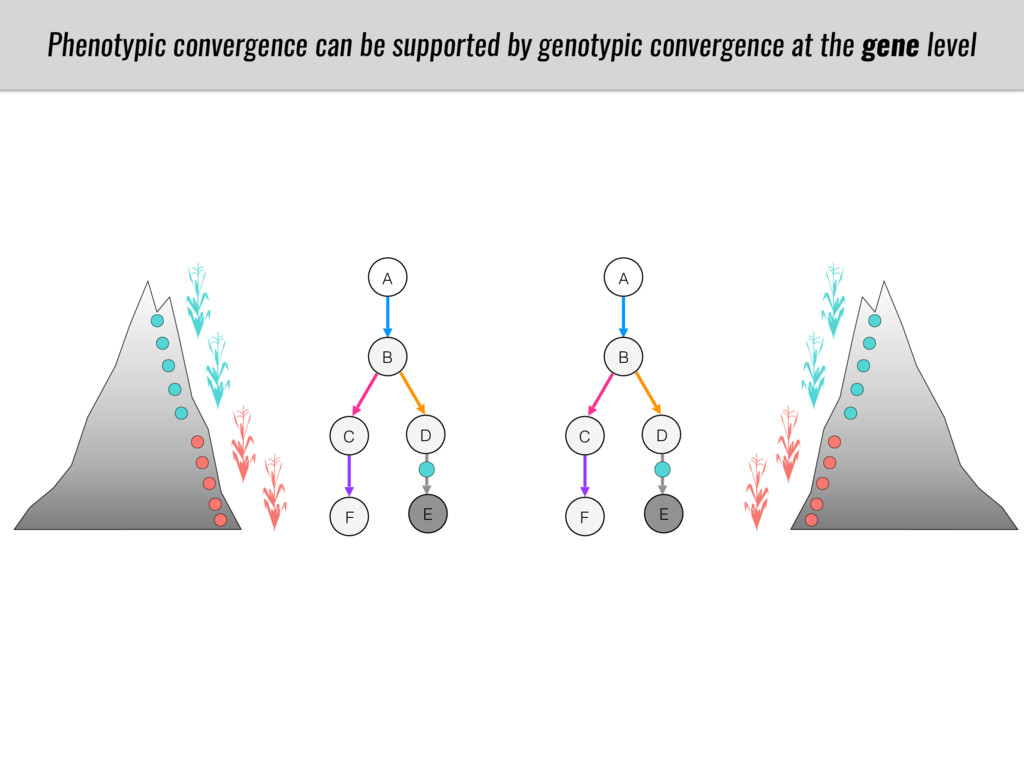
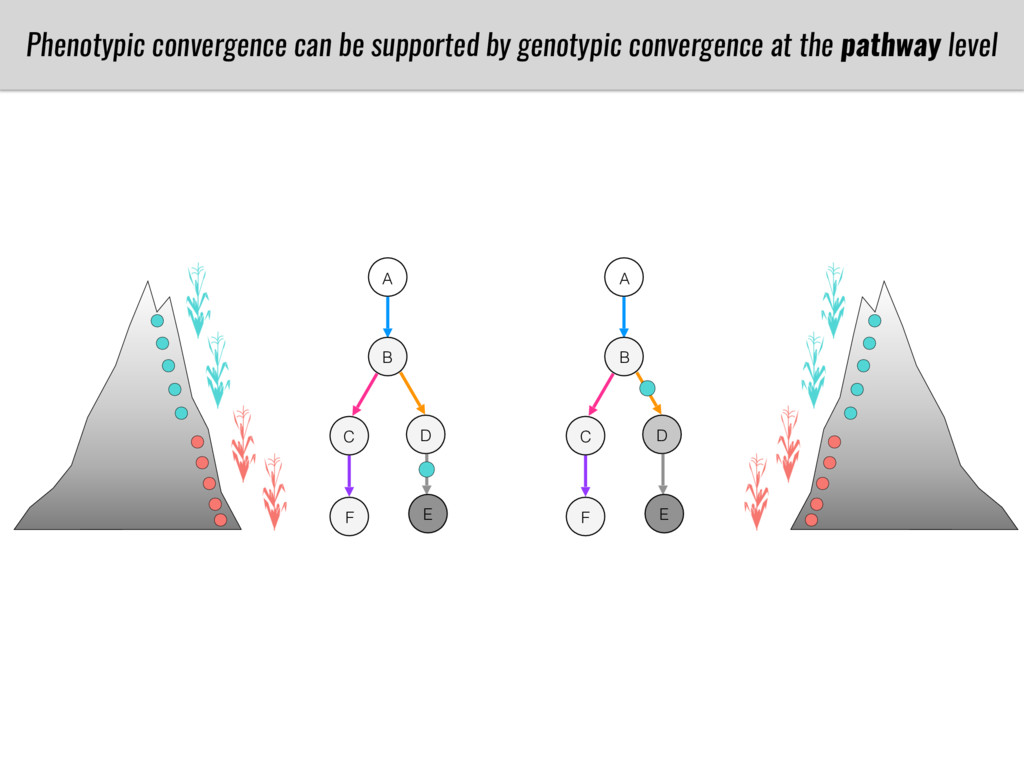
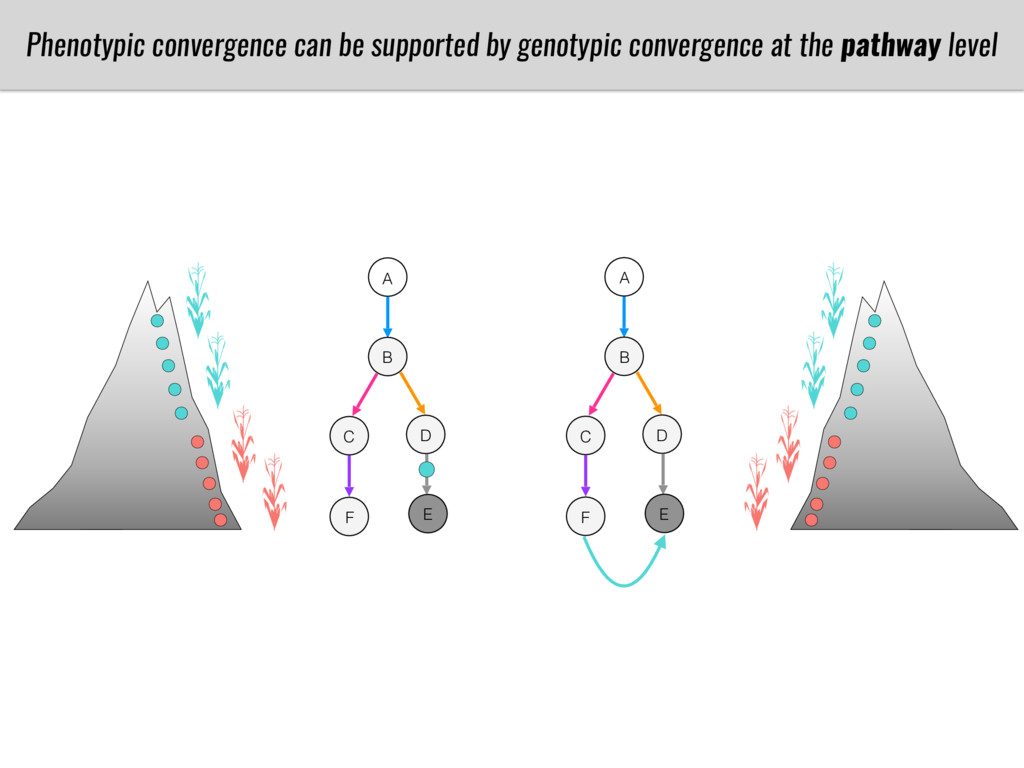
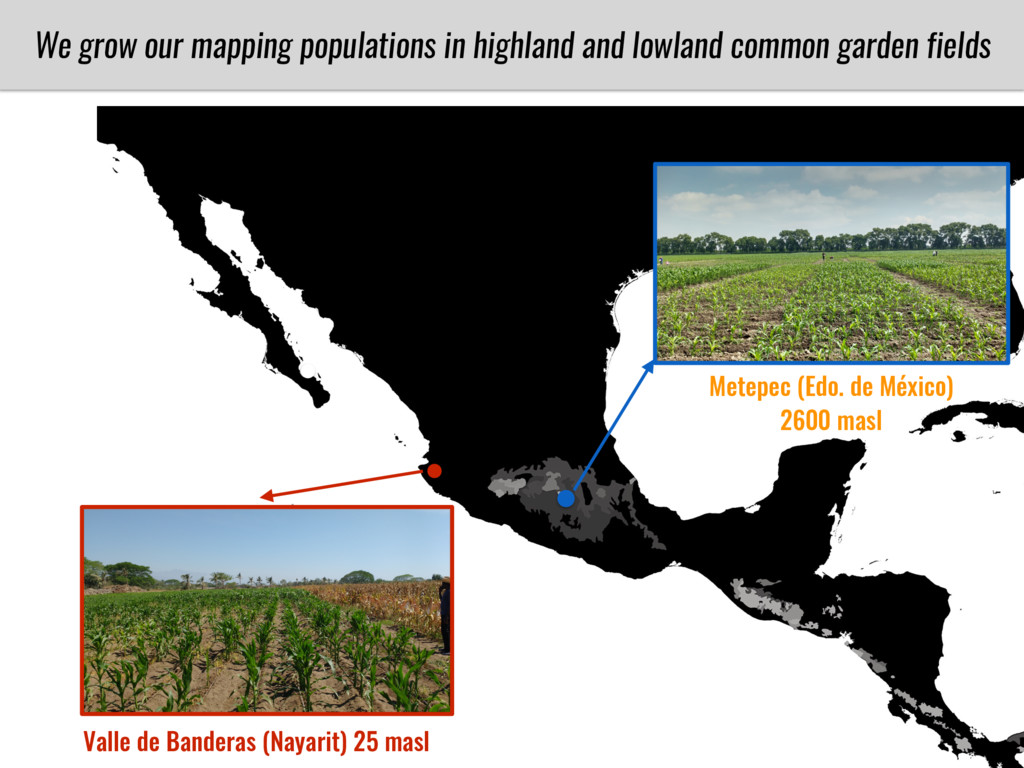
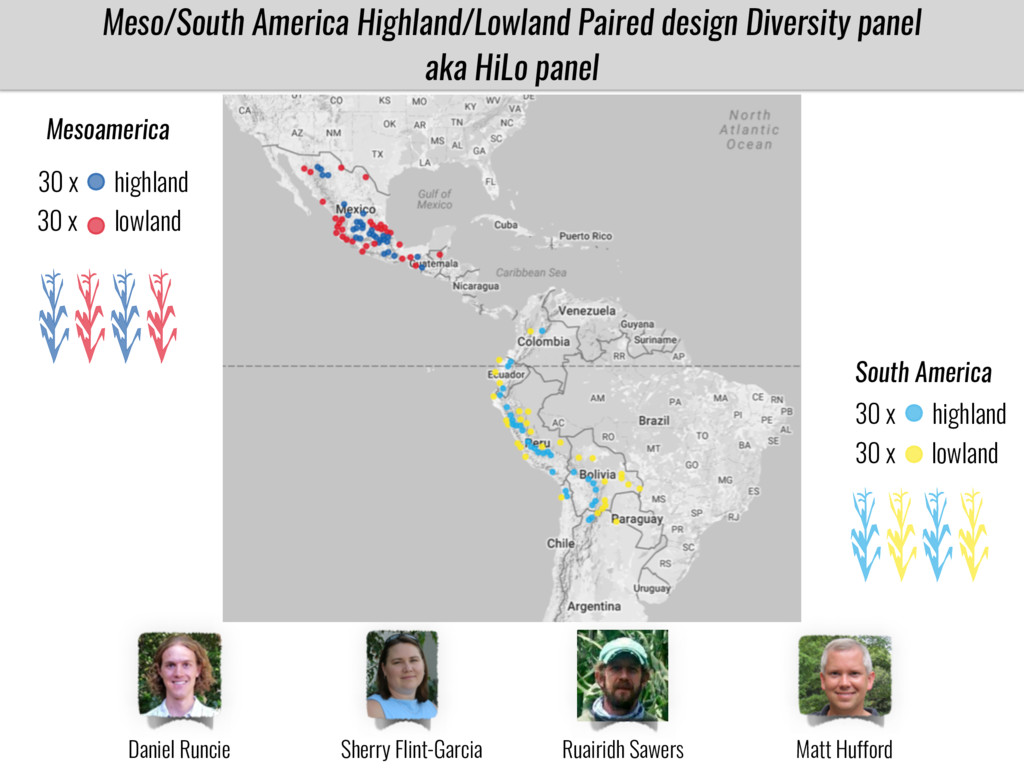
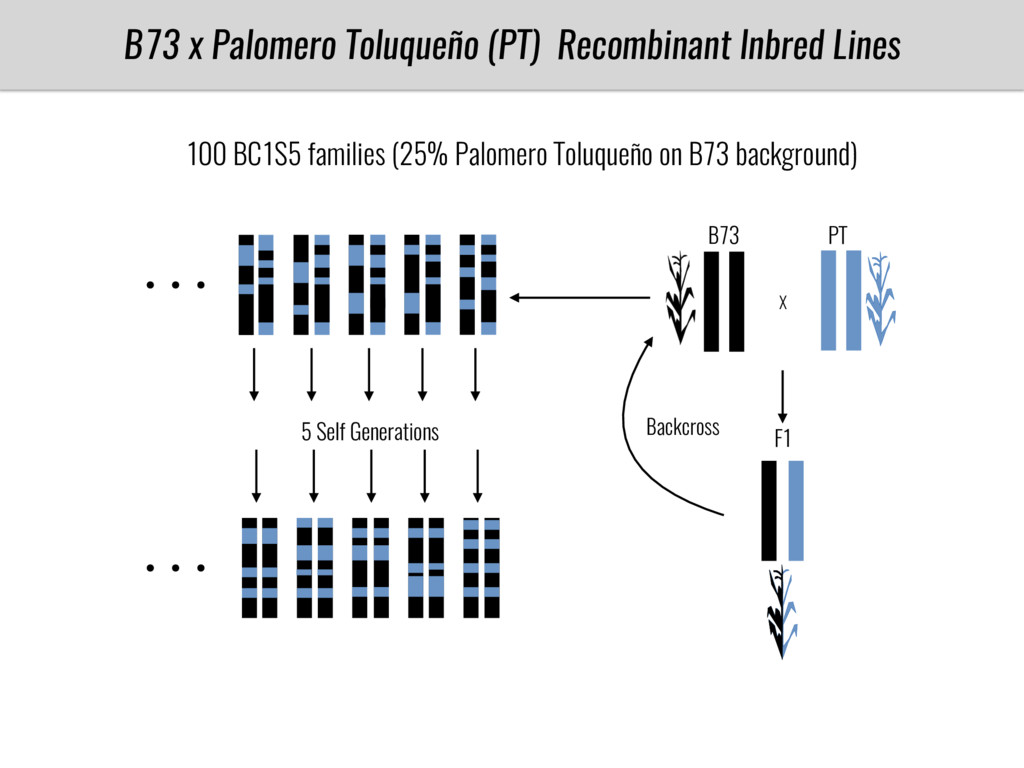
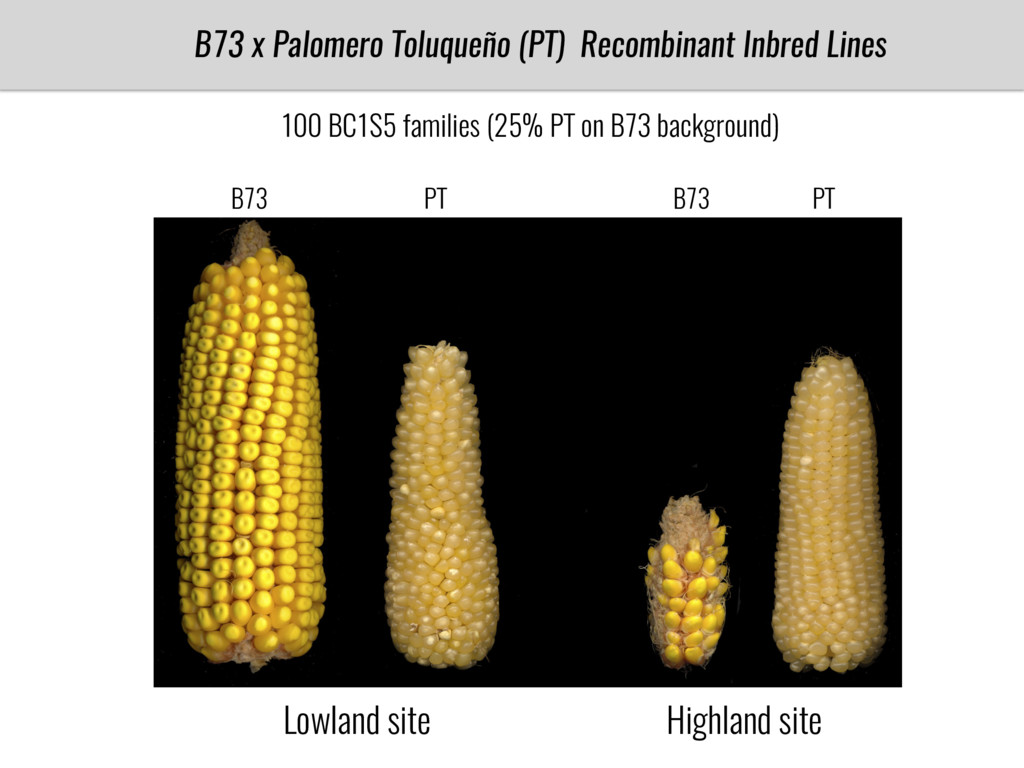
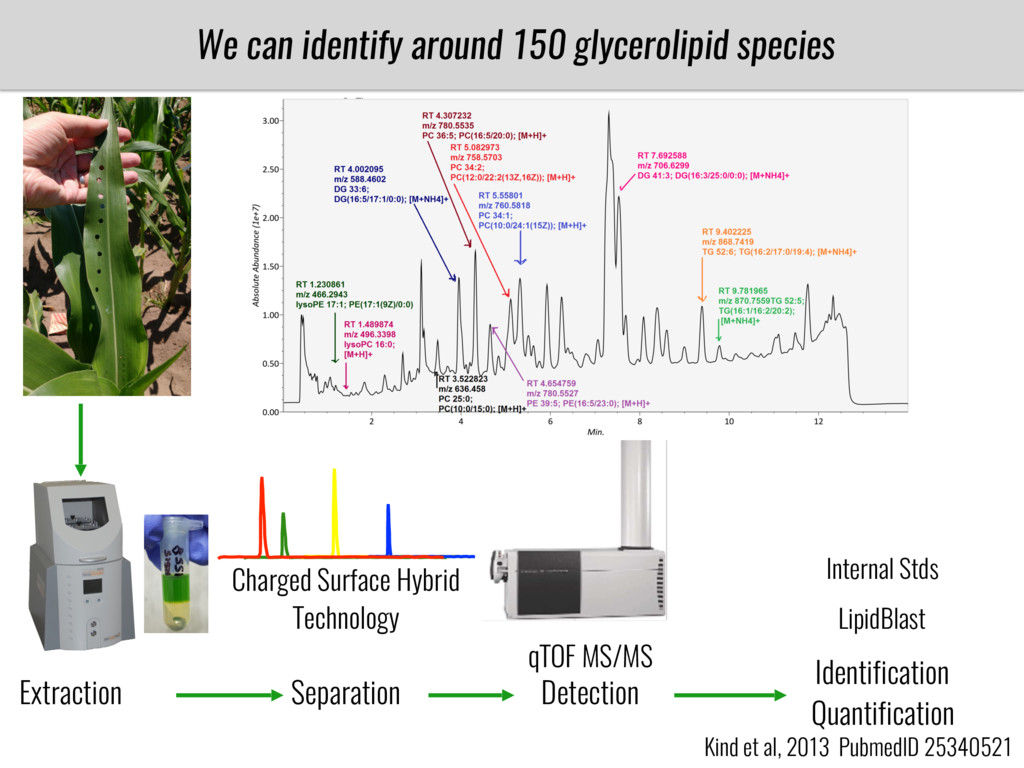
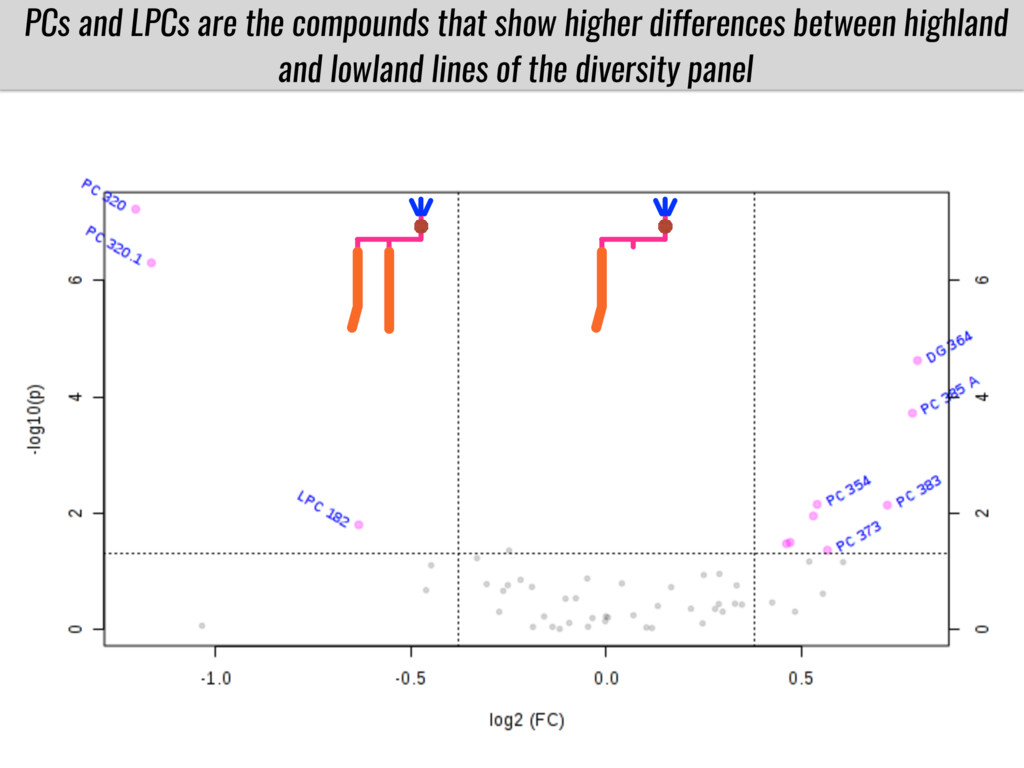
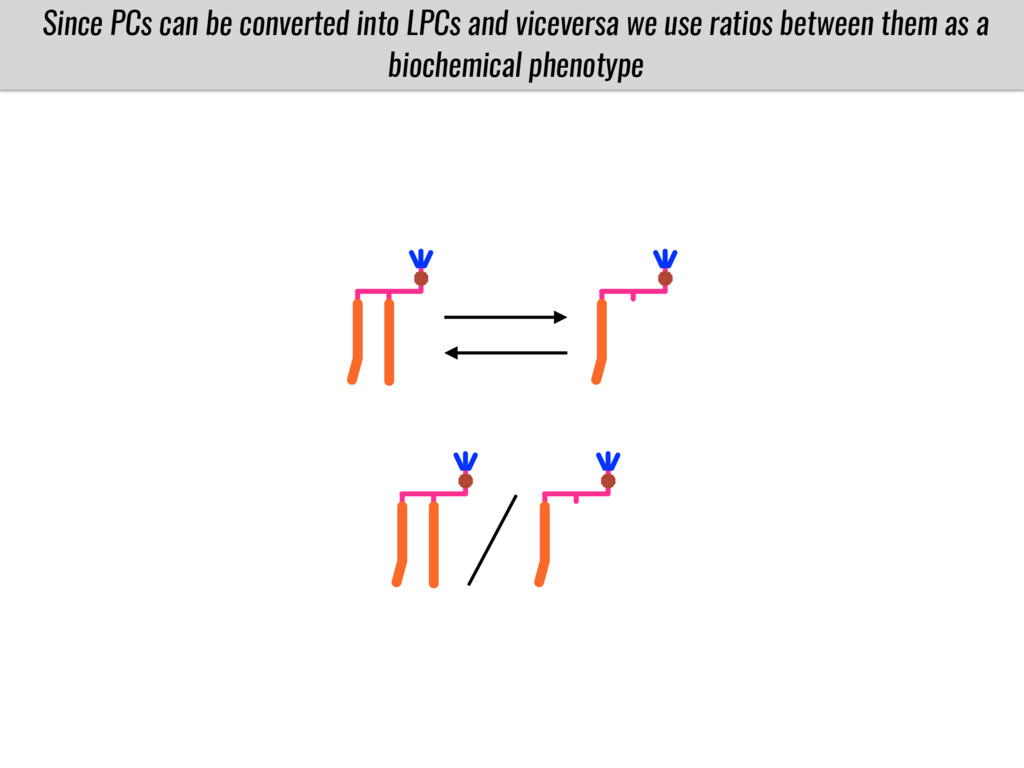
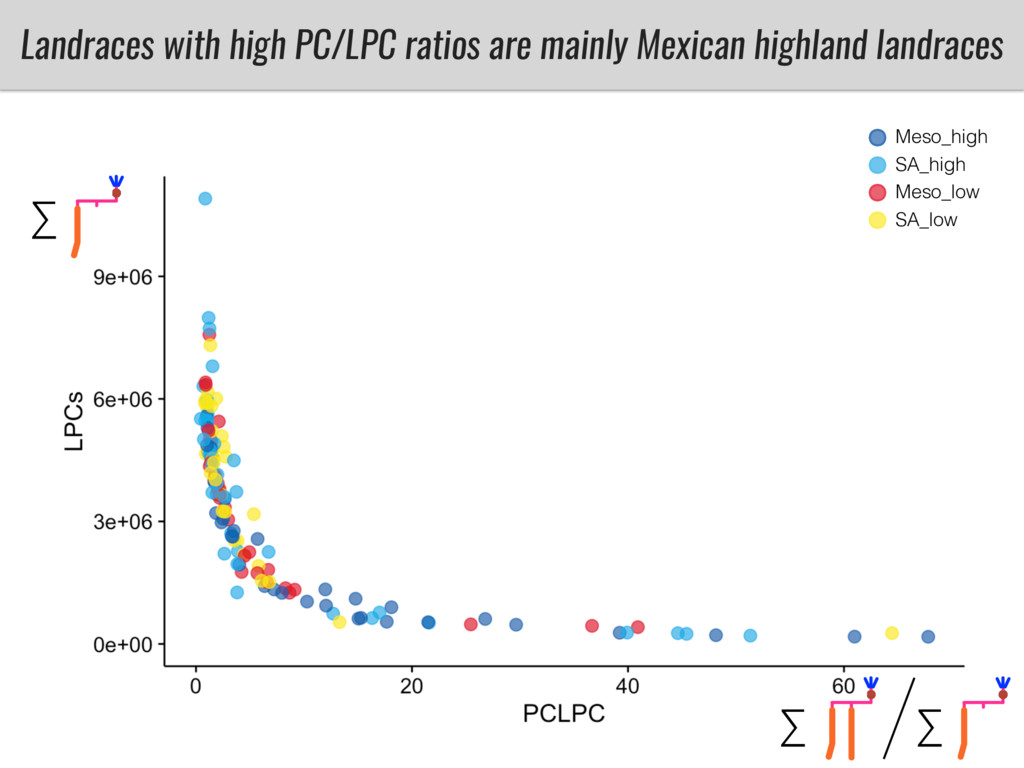
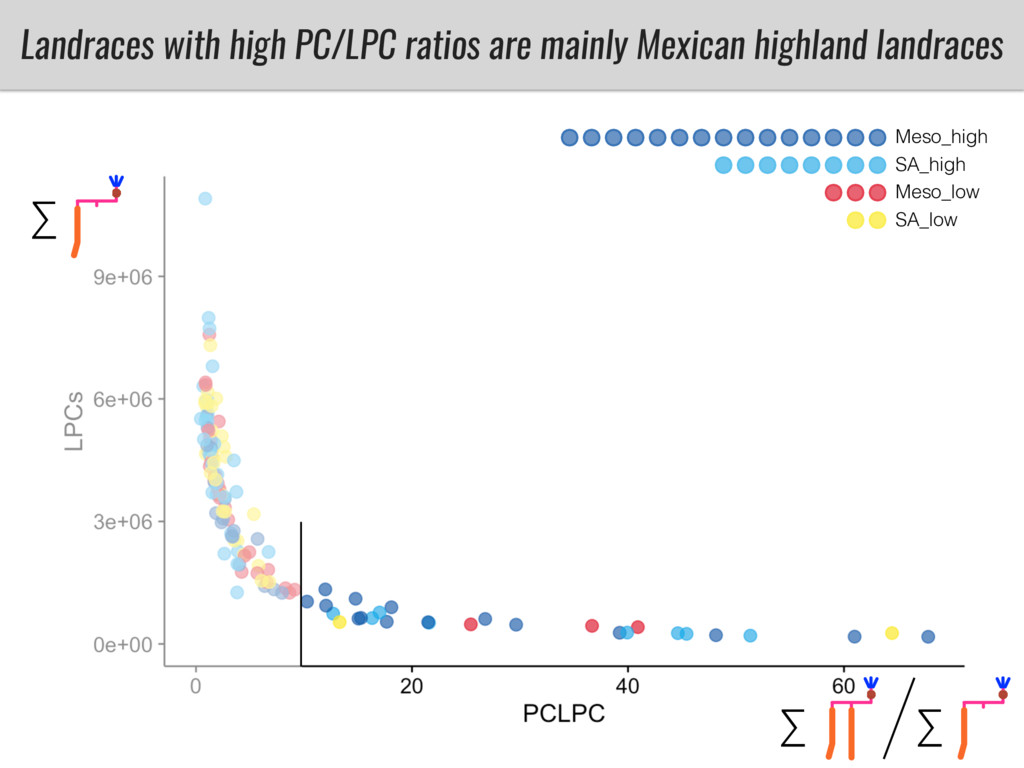
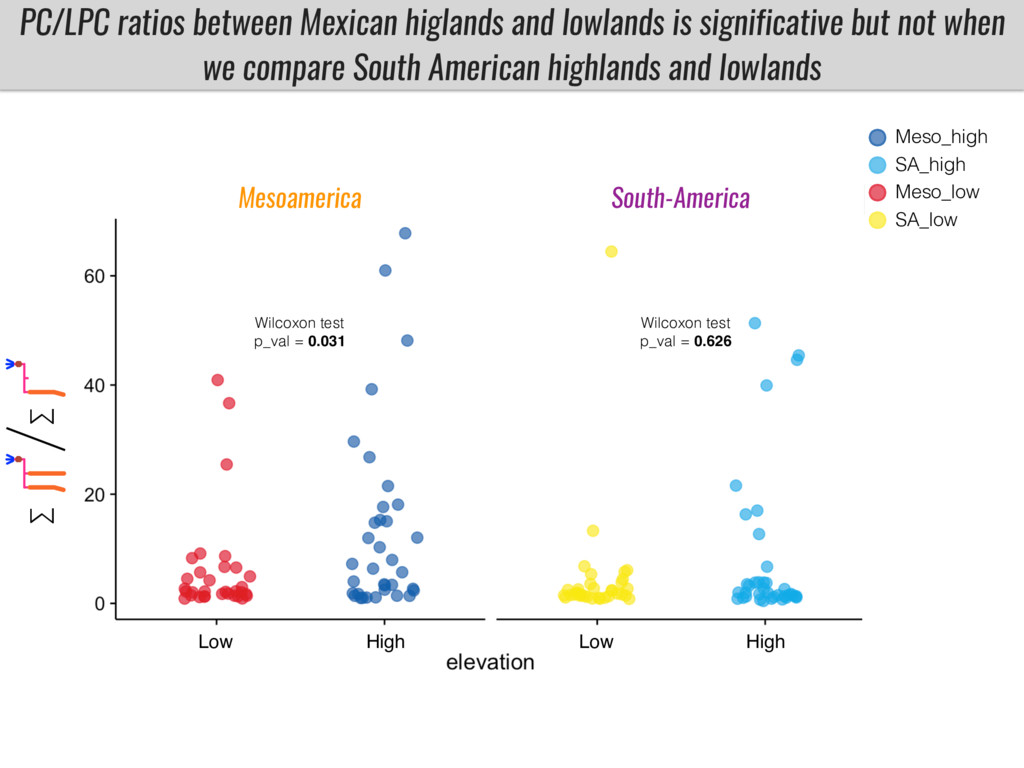
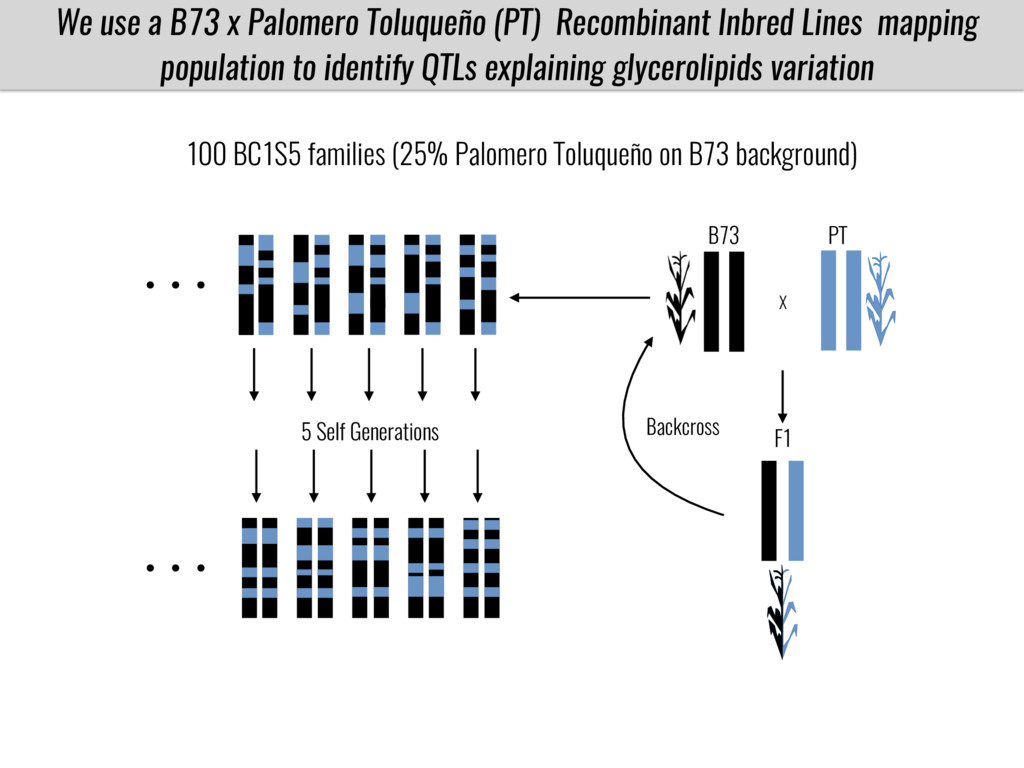
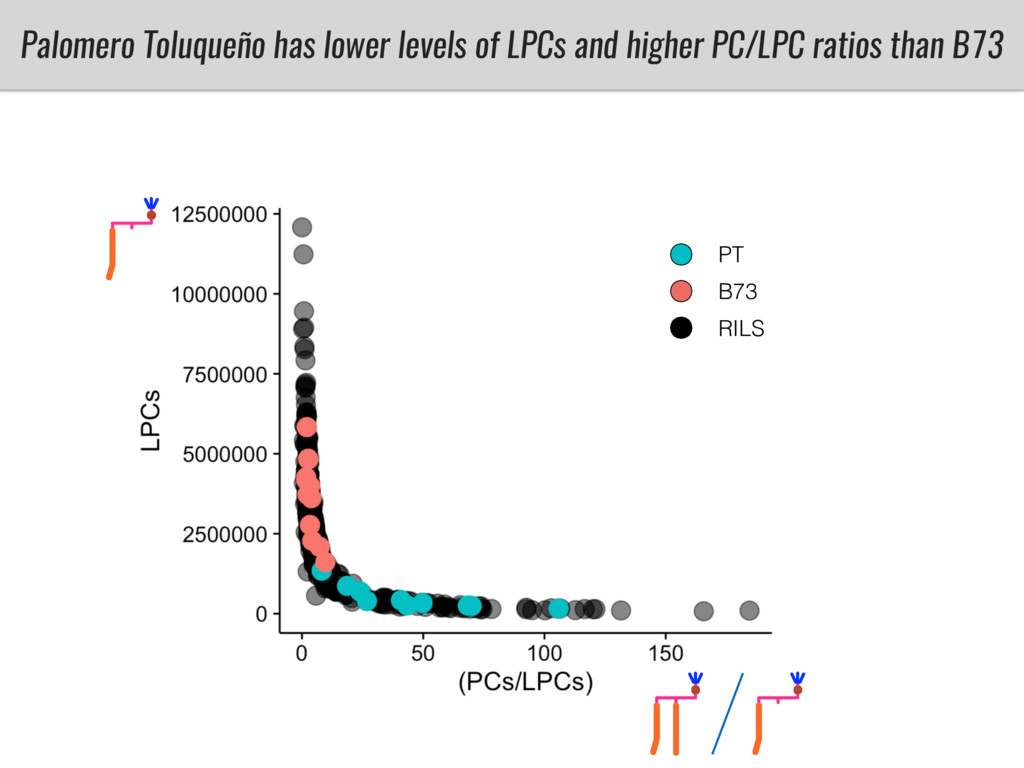
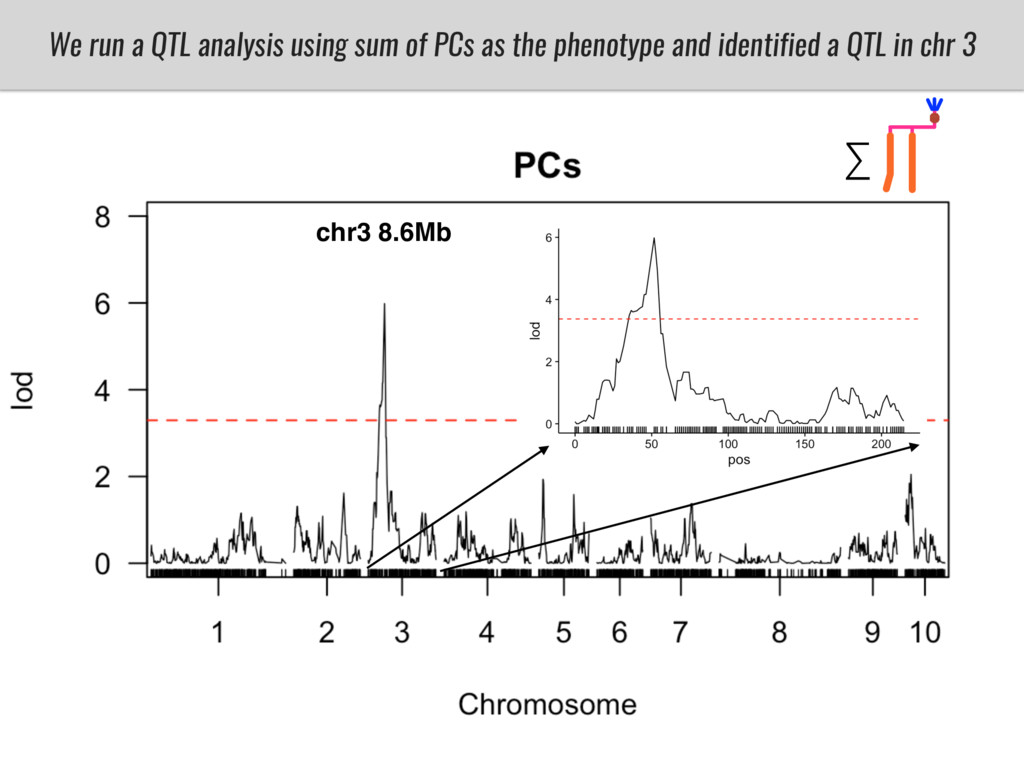
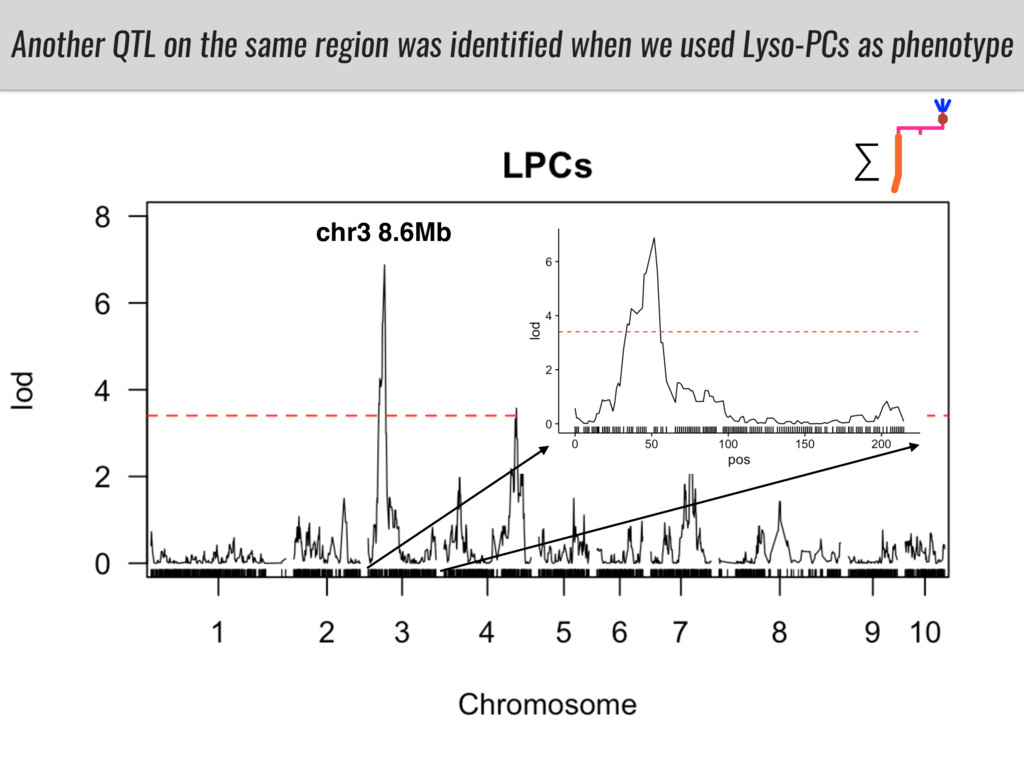
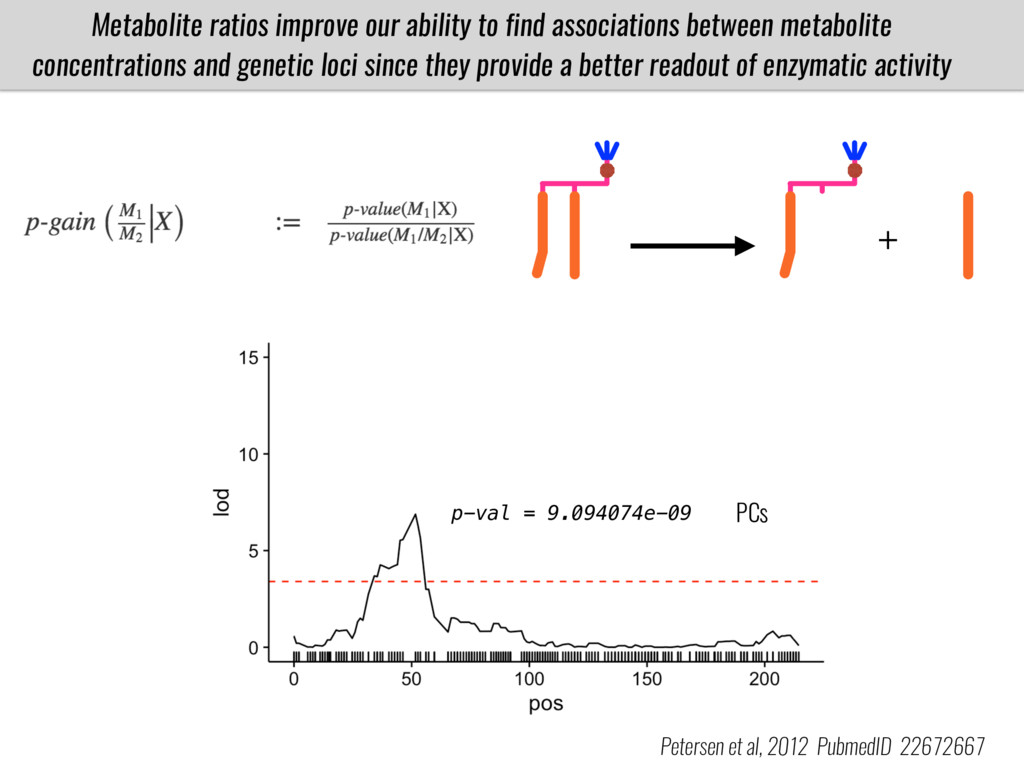
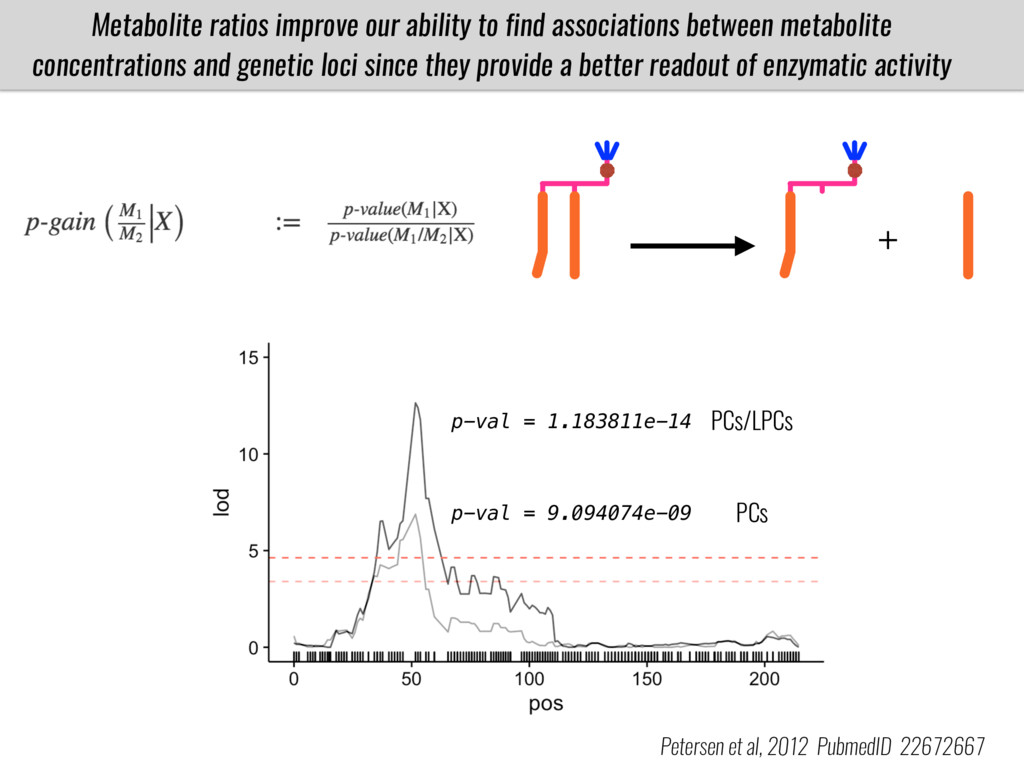
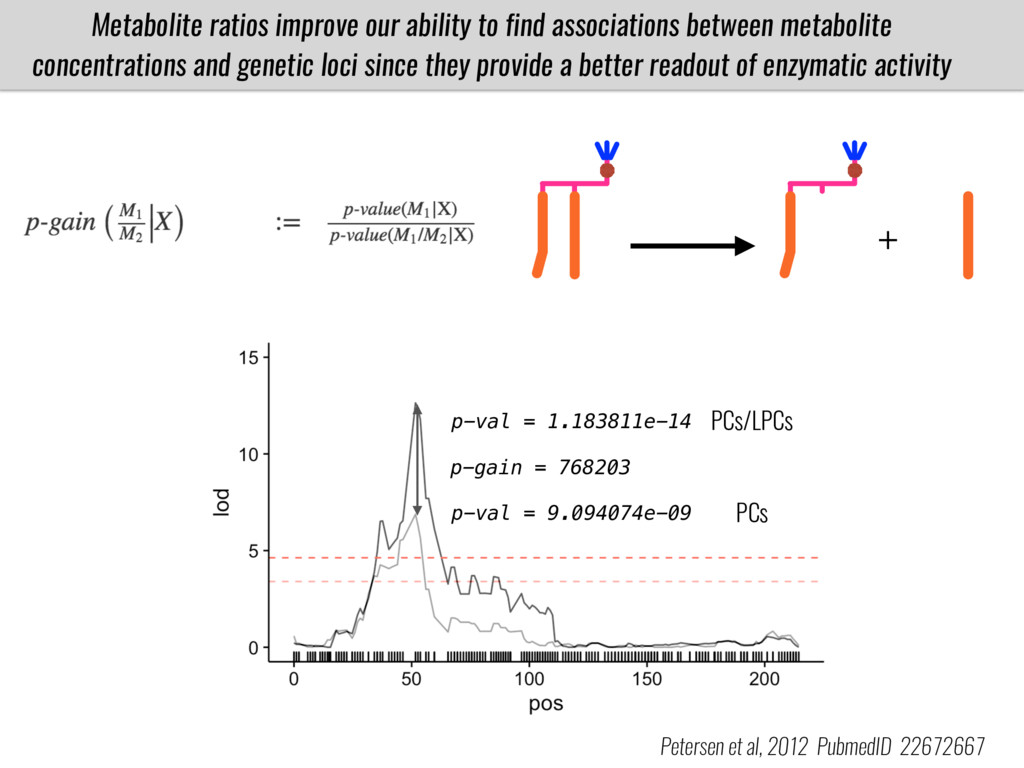
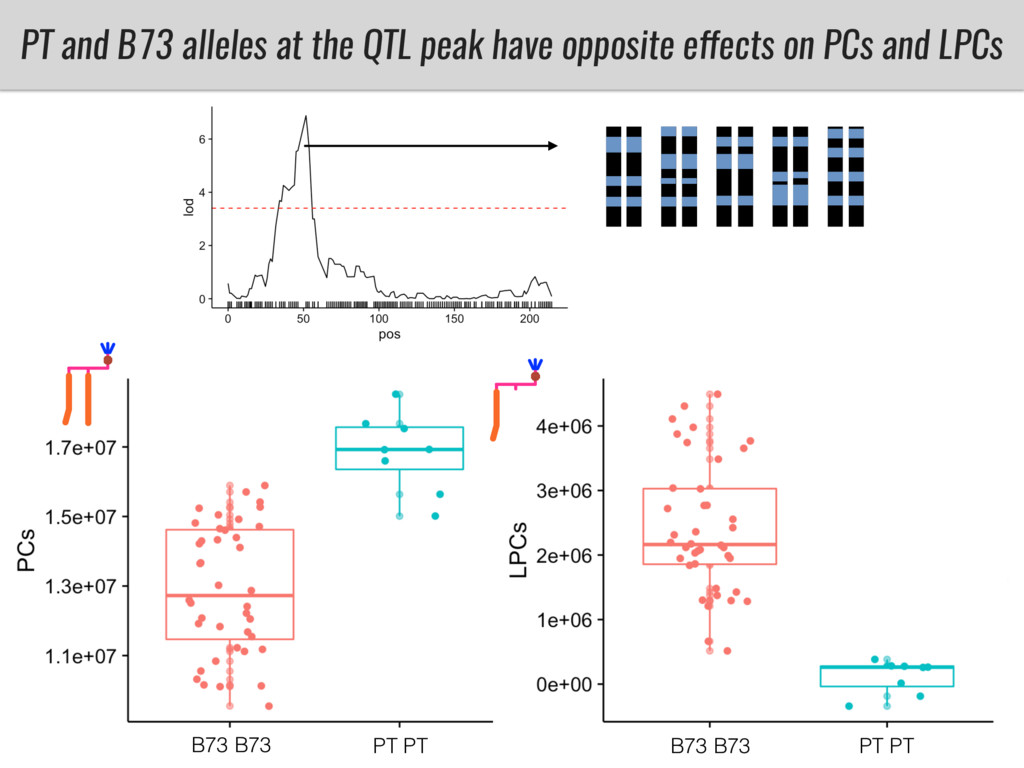
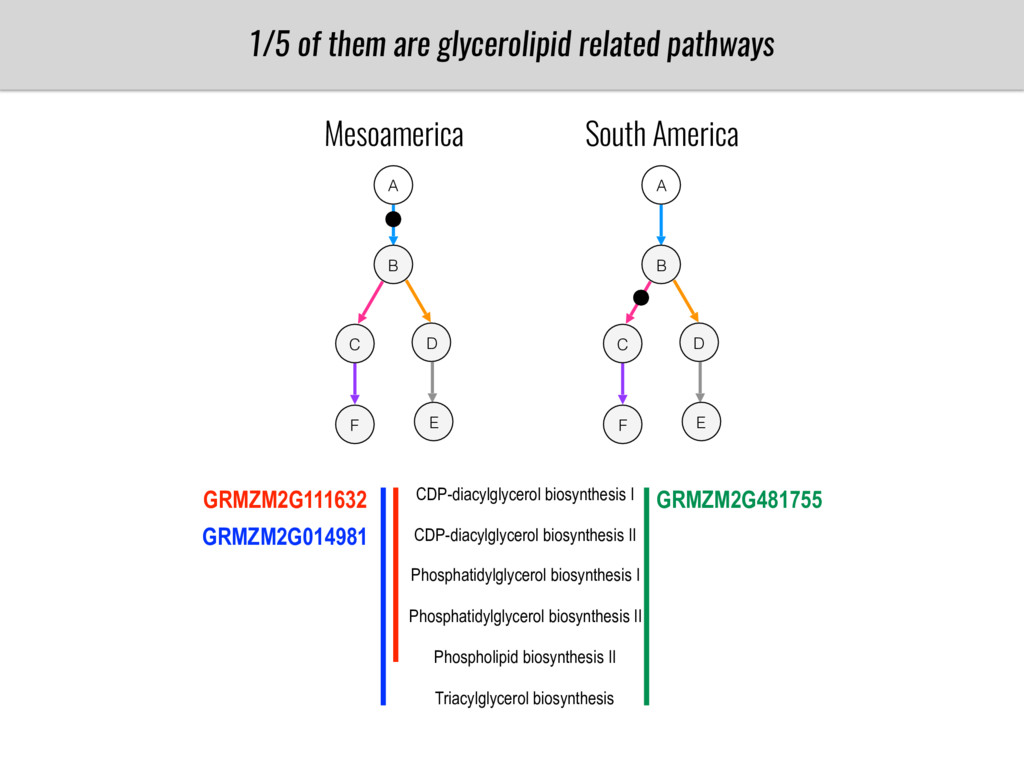
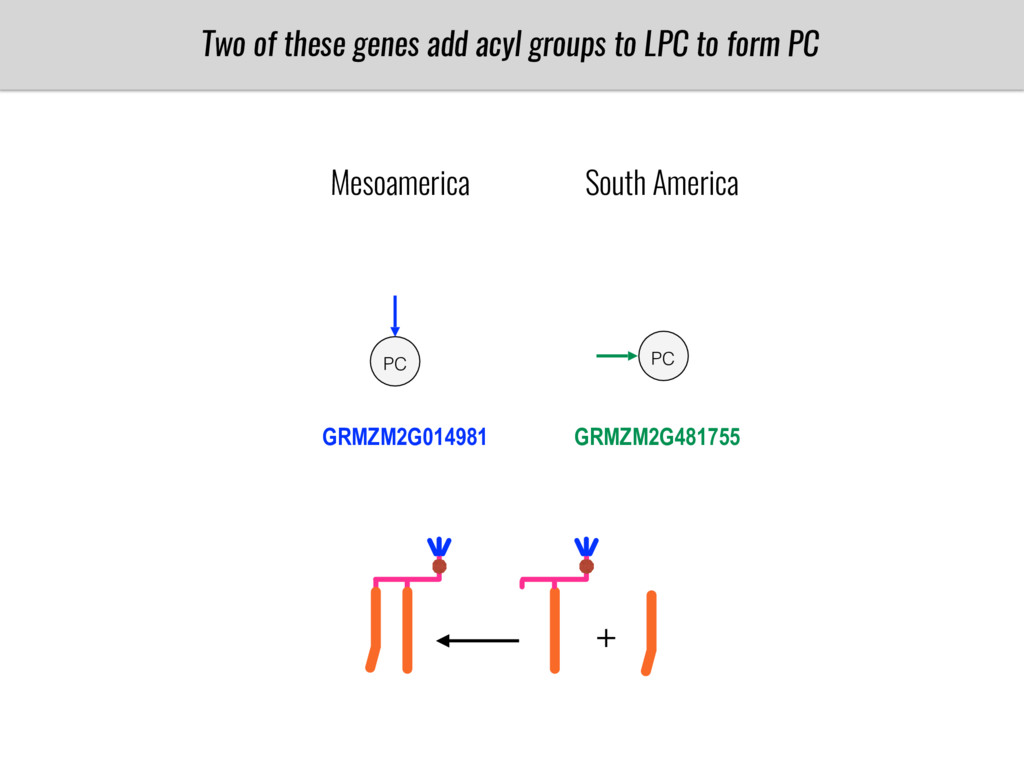
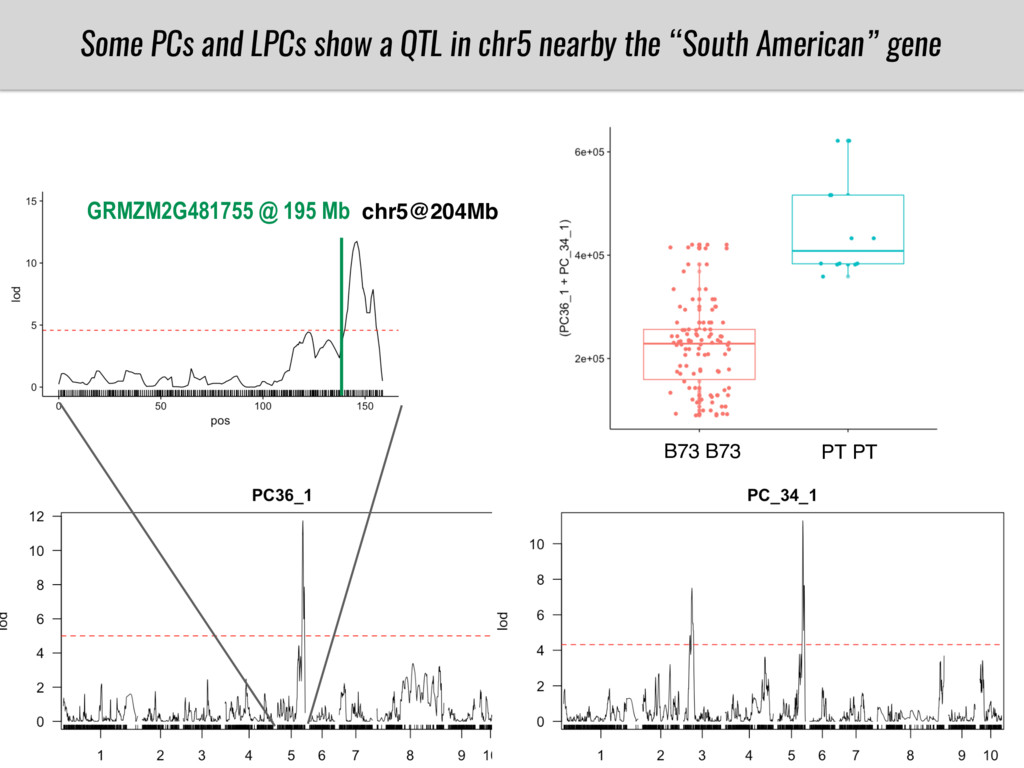
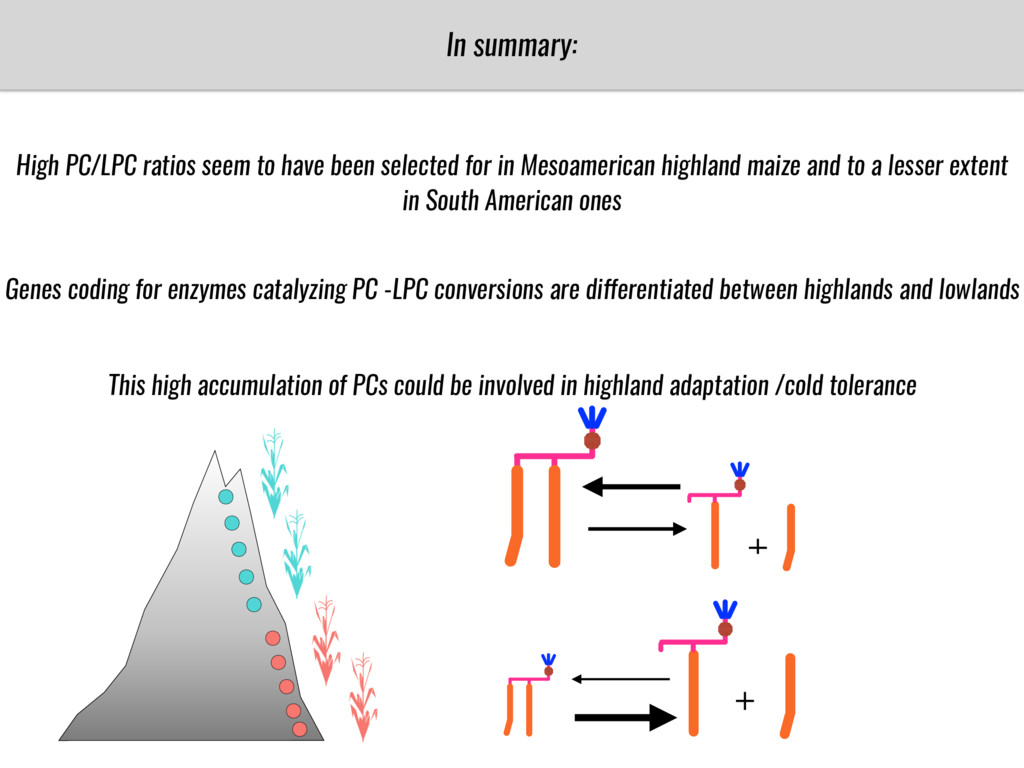
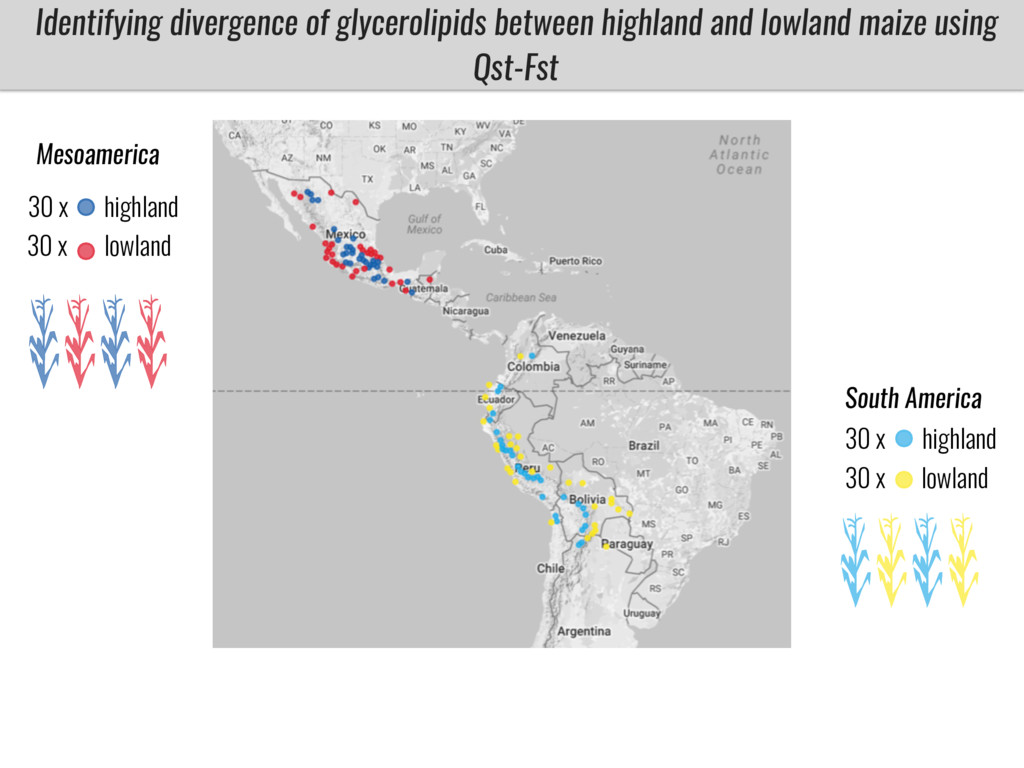
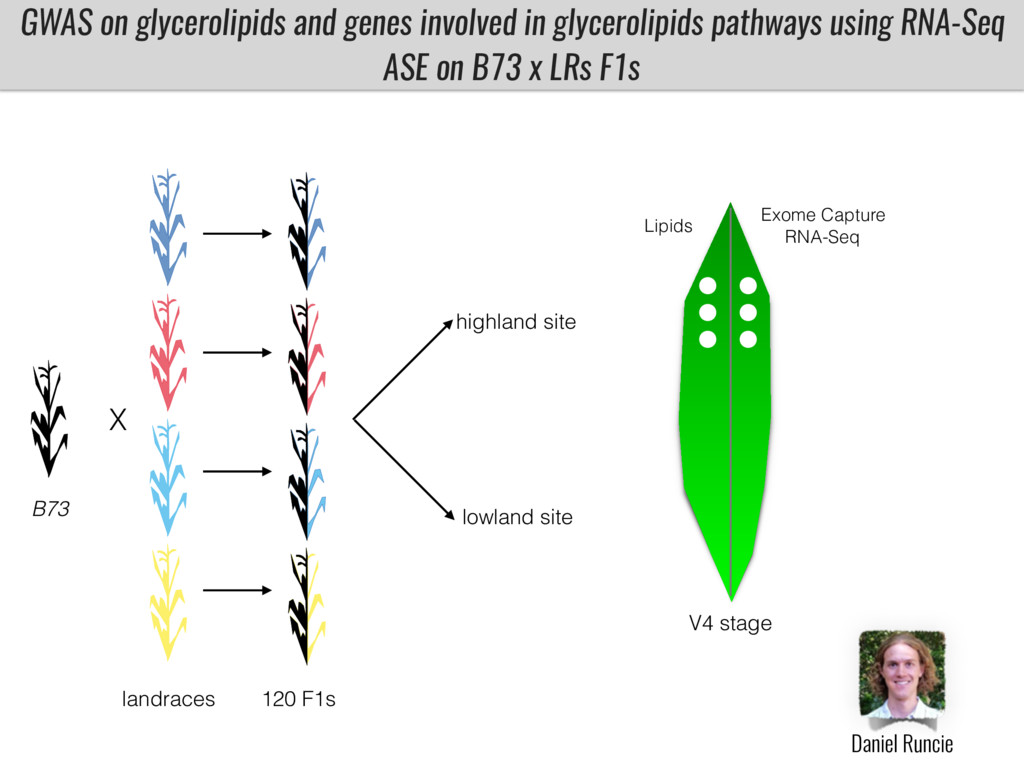
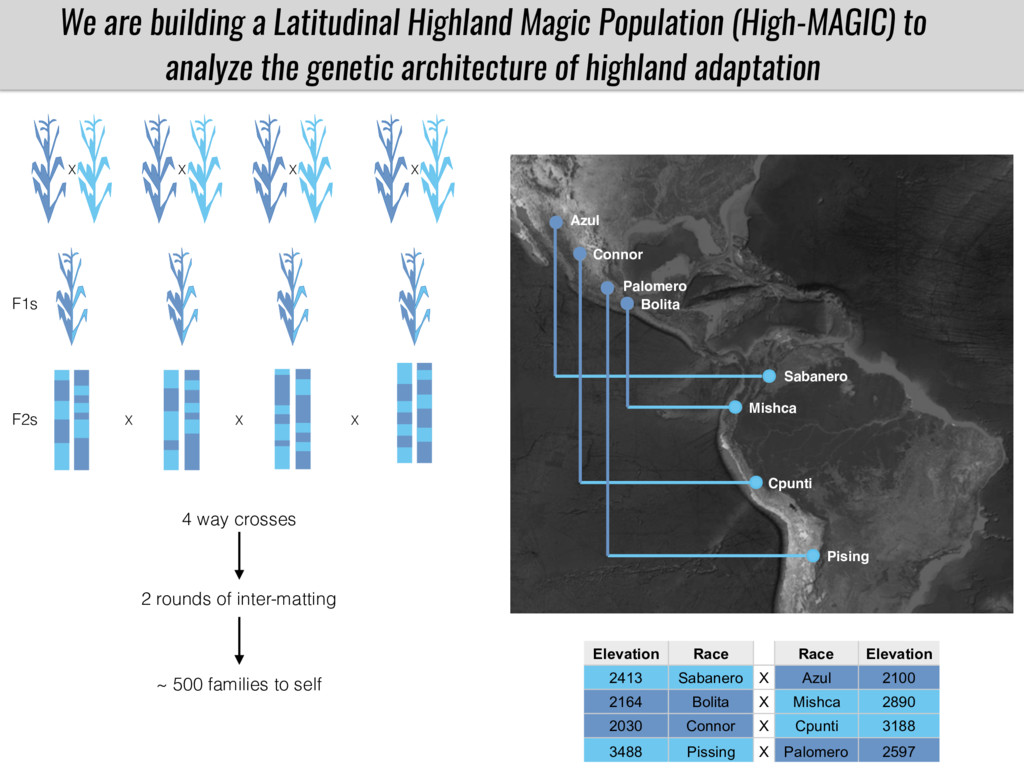
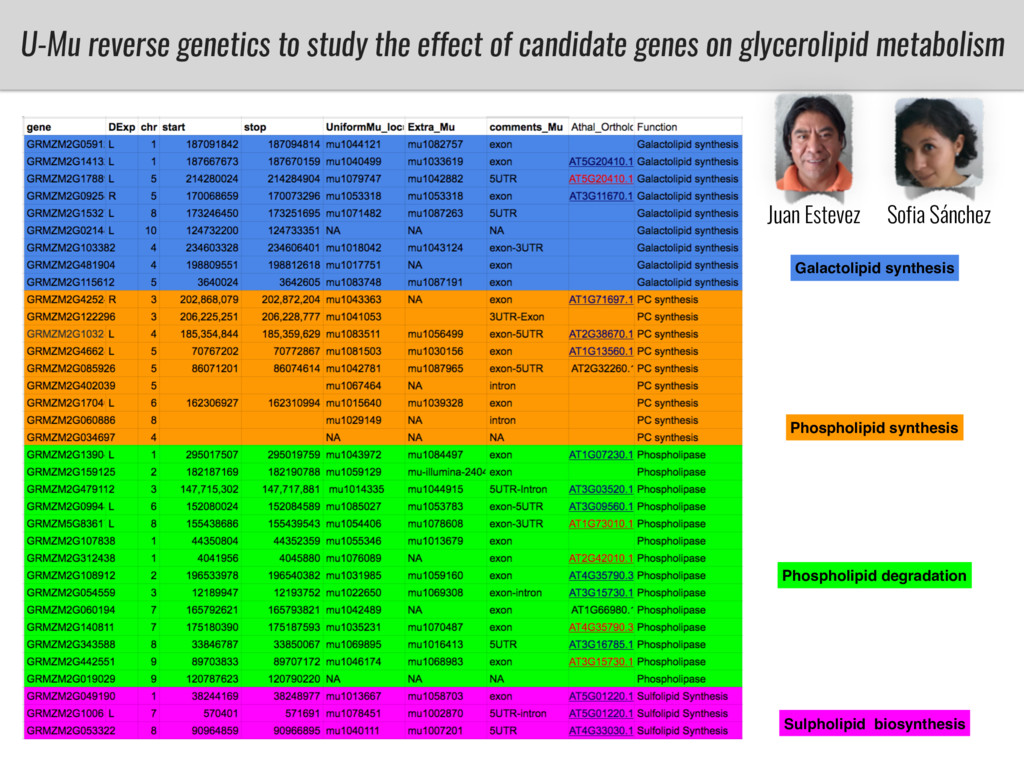
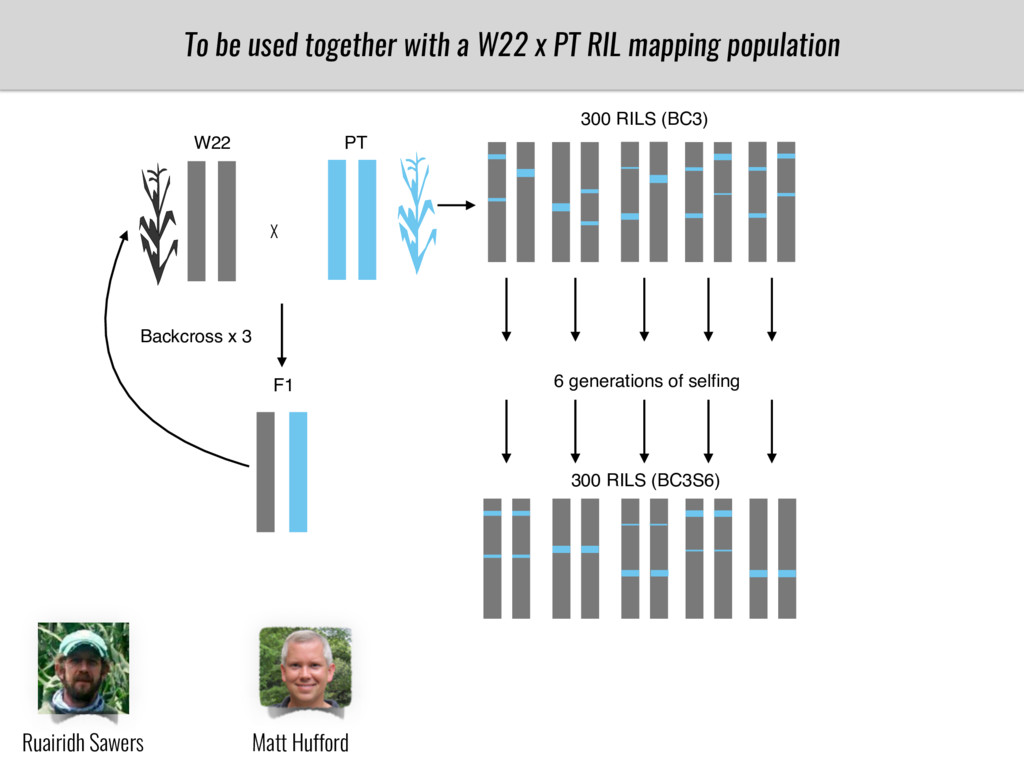
To test this hypothesis grew a maize RIL mapping population (developed using a Mexican highland landrace Palomero Toluqueño and B73) and a 120 landrace diversity panel (composed of lowland and highland Mesoamerican and South American landraces) in common garden experiments at sea level and at 2600 masl. in Mexico and measures glycerolipid species (125 different compounds) that were used as phenotypes to identify genotype-phenotype associations and plasticity between highland and lowland conditions. I will present this results and in particular on an interesting QTL in ch3 that explains phosphatidylcholine to lysophosphatidylcholine conversion and seems to be selected against in highland Mexican landraces but not in South American ones.
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
![3500 masl ~ 11500 feet pH ~ 5.1 [P] ~](https://files.speakerdeck.com/presentations/ce83cde51ea649048e78cdf04b8f28b5/slide_38.jpg){kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}