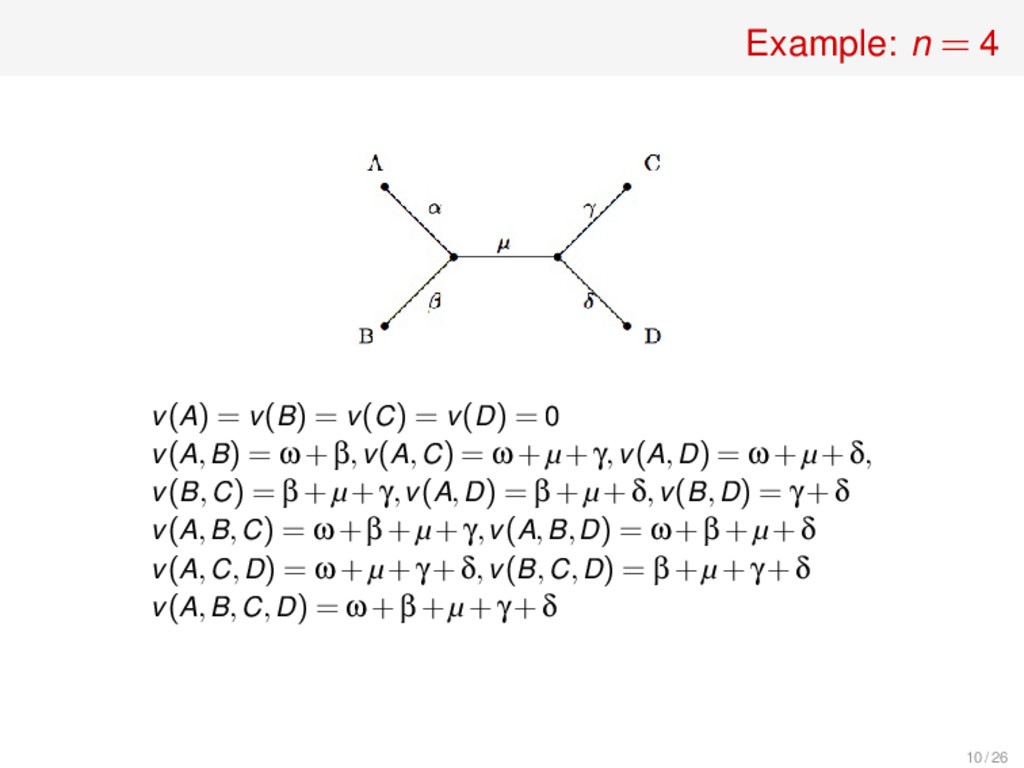
v(D) = 0 v(A,B) = ω+β,v(A,C) = ω+µ+γ,v(A,D) = ω+µ+δ, v(B,C) = β+µ+γ,v(A,D) = β+µ+δ,v(B,D) = γ+δ v(A,B,C) = ω+β+µ+γ,v(A,B,D) = ω+β+µ+δ v(A,C,D) = ω+µ+γ+δ,v(B,C,D) = β+µ+γ+δ v(A,B,C,D) = ω+β+µ+γ+δ ϕ(A) = 3! 4! (v(A)−v(/ 0)) +2! 4! ((v(A,B)−v(B))+(v(A,C)−v(C))+(v(A,D)−v(D))) +2! 4! ((v(A,B,C)−v(B,C))+(v(A,B,D)−v(B,D)) +(v(A,C,D)−v(C,D))) +3! 4! (v(A,B,C,D)−v(B,C,D)) = 1 4 ·0 + 1 12 (3ω+2µ+β+γ+δ)+ 1 12 (3ω+µ)+ 1 4 ω = 9 12 ω+ 1 12 β+ 1 12 γ+ 1 12 δ+ 3 12 µ 11 / 26
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}