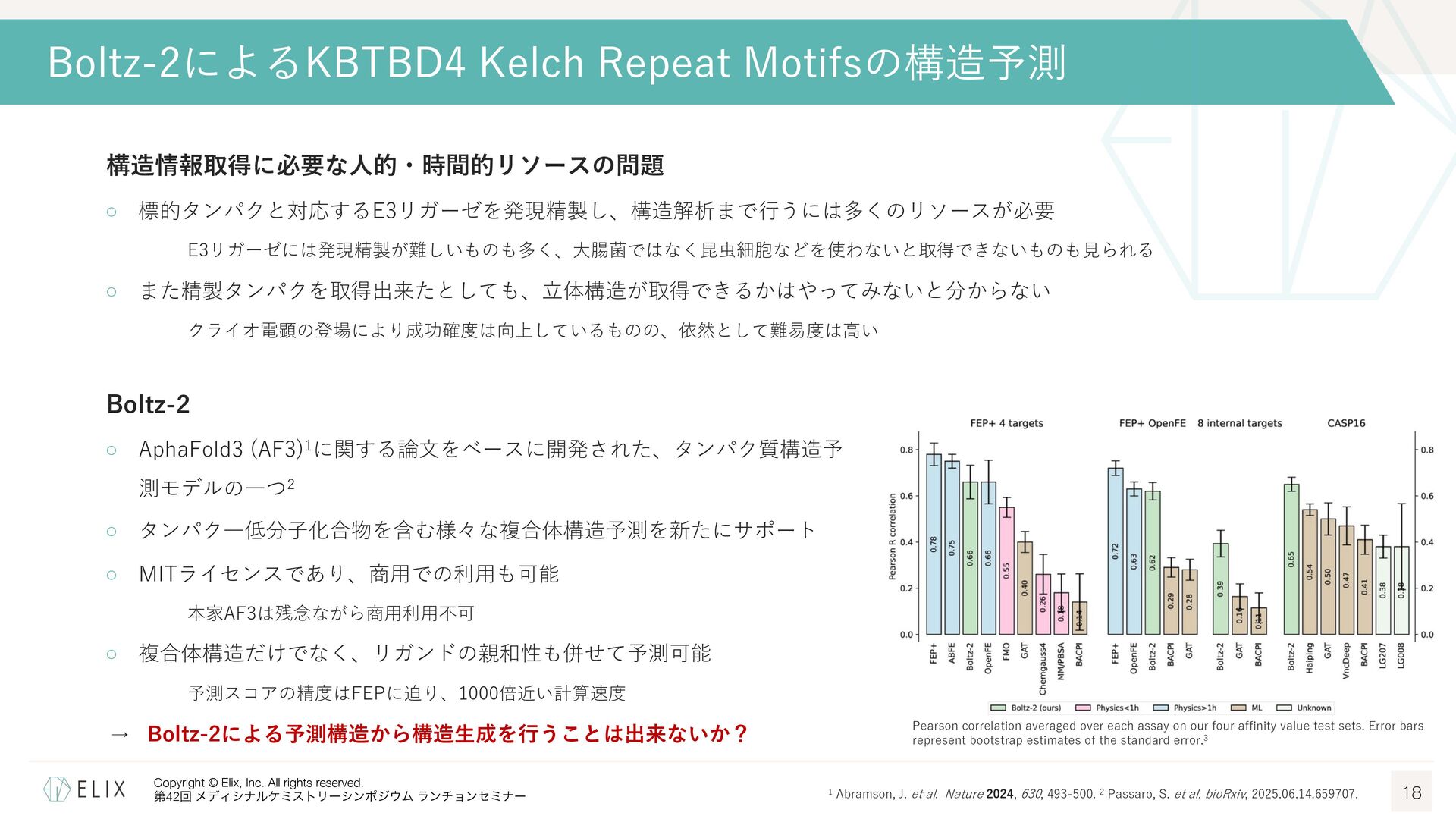
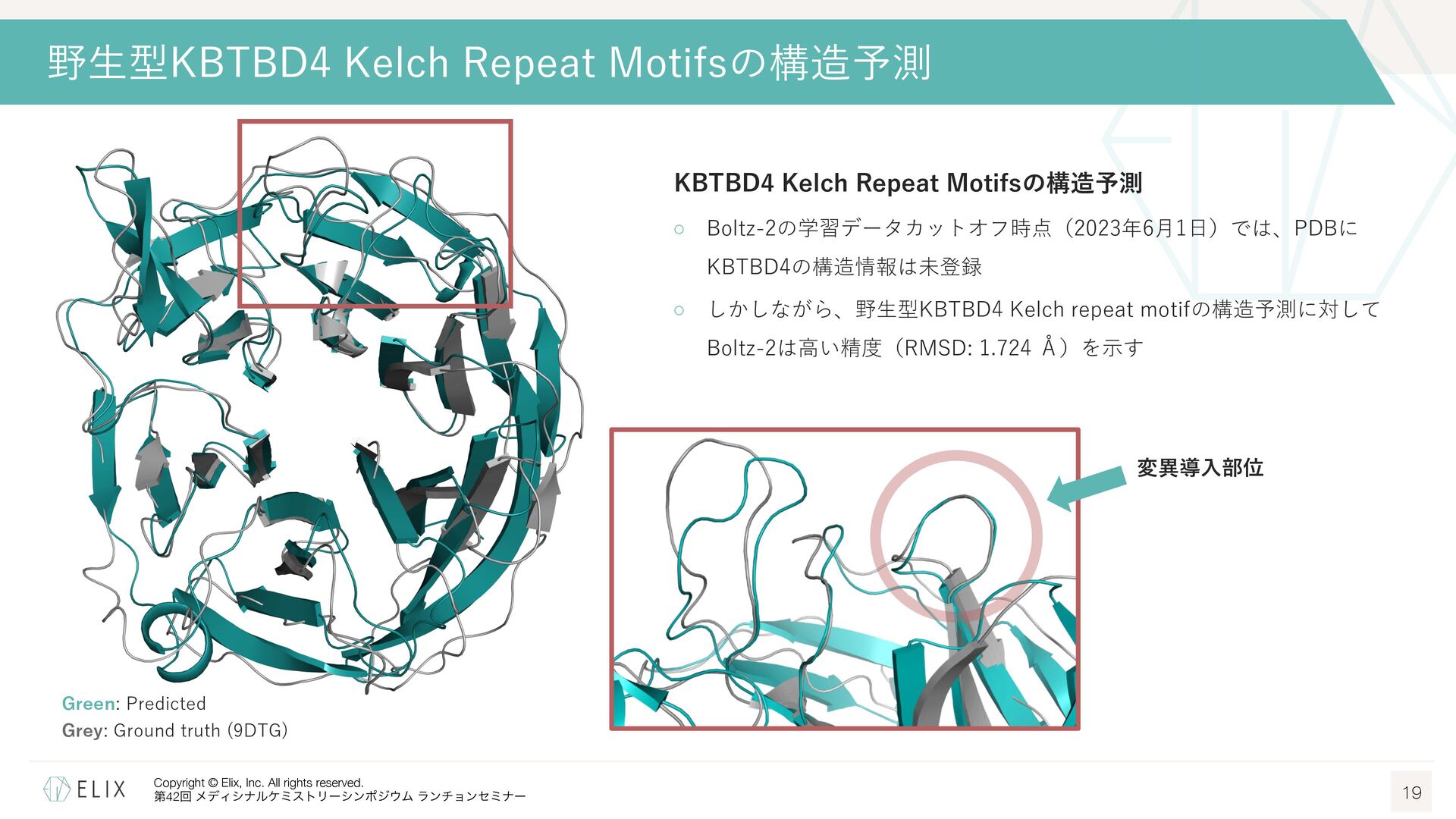
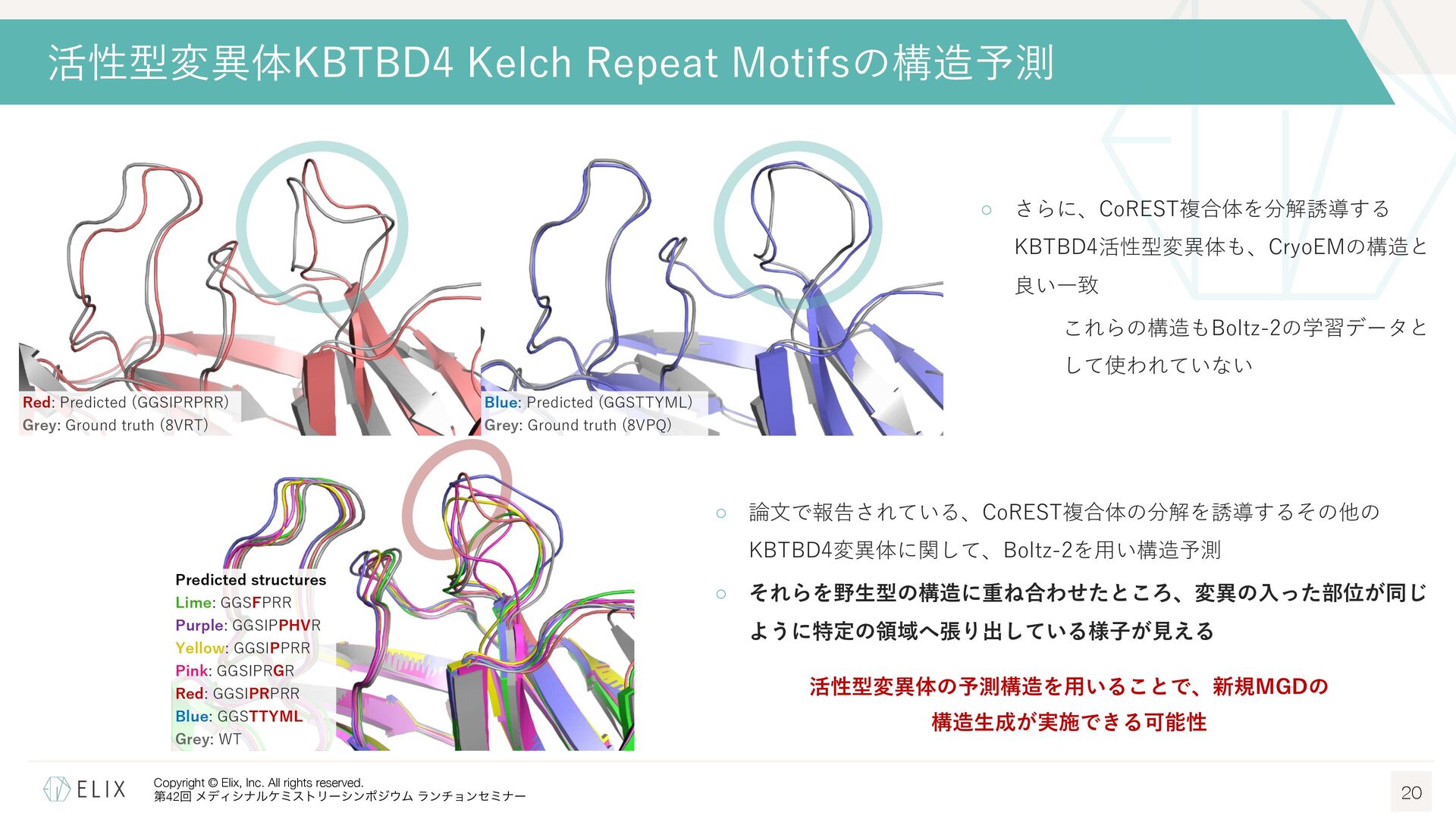
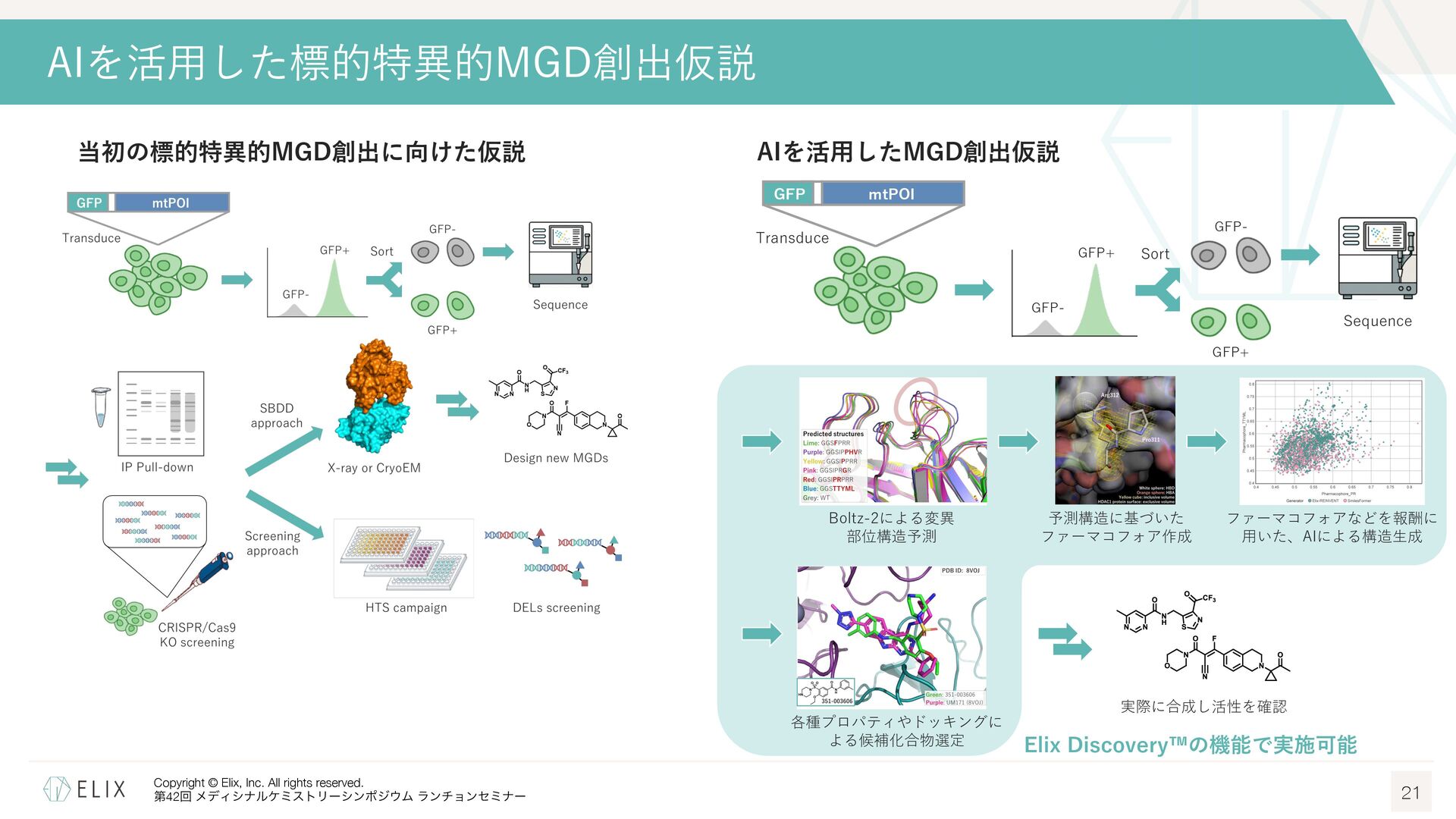
構造情報取得に必要な⼈的・時間的リソースの問題 ◦ 標的タンパクと対応するE3リガーゼを発現精製し、構造解析まで⾏うには多くのリソースが必要 E3リガーゼには発現精製が難しいものも多く、⼤腸菌ではなく昆⾍細胞などを使わないと取得できないものも⾒られる ◦ また精製タンパクを取得出来たとしても、⽴体構造が取得できるかはやってみないと分からない クライオ電顕の登場により成功確度は向上しているものの、依然として難易度は⾼い Boltz-2によるKBTBD4 Kelch Repeat Motifsの構造予測 18 Boltz-2 ◦ AphaFold3 (AF3)1に関する論⽂をベースに開発された、タンパク質構造予 測モデルの⼀つ2 ◦ タンパク―低分⼦化合物を含む様々な複合体構造予測を新たにサポート ◦ MITライセンスであり、商⽤での利⽤も可能 本家AF3は残念ながら商⽤利⽤不可 ◦ 複合体構造だけでなく、リガンドの親和性も併せて予測可能 予測スコアの精度はFEPに迫り、1000倍近い計算速度 → Boltz-2による予測構造から構造⽣成を⾏うことは出来ないか? Pearson correlation averaged over each assay on our four affinity value test sets. Error bars represent bootstrap estimates of the standard error.3 1 Abramson, J. et al. Nature 2024, 630, 493-500. 2 Passaro, S. et al. bioRxiv, 2025.06.14.659707.
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
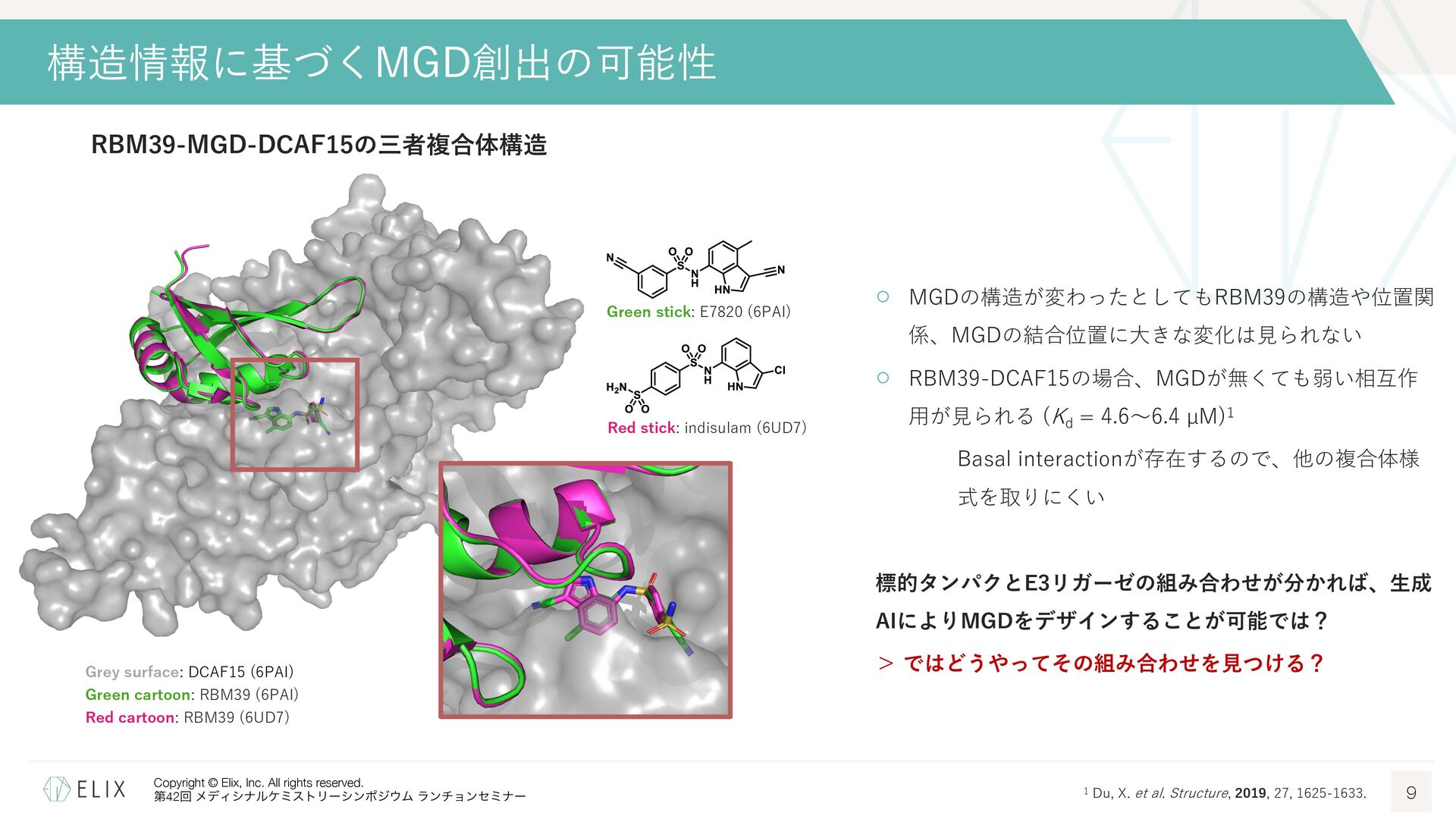{kind=link}
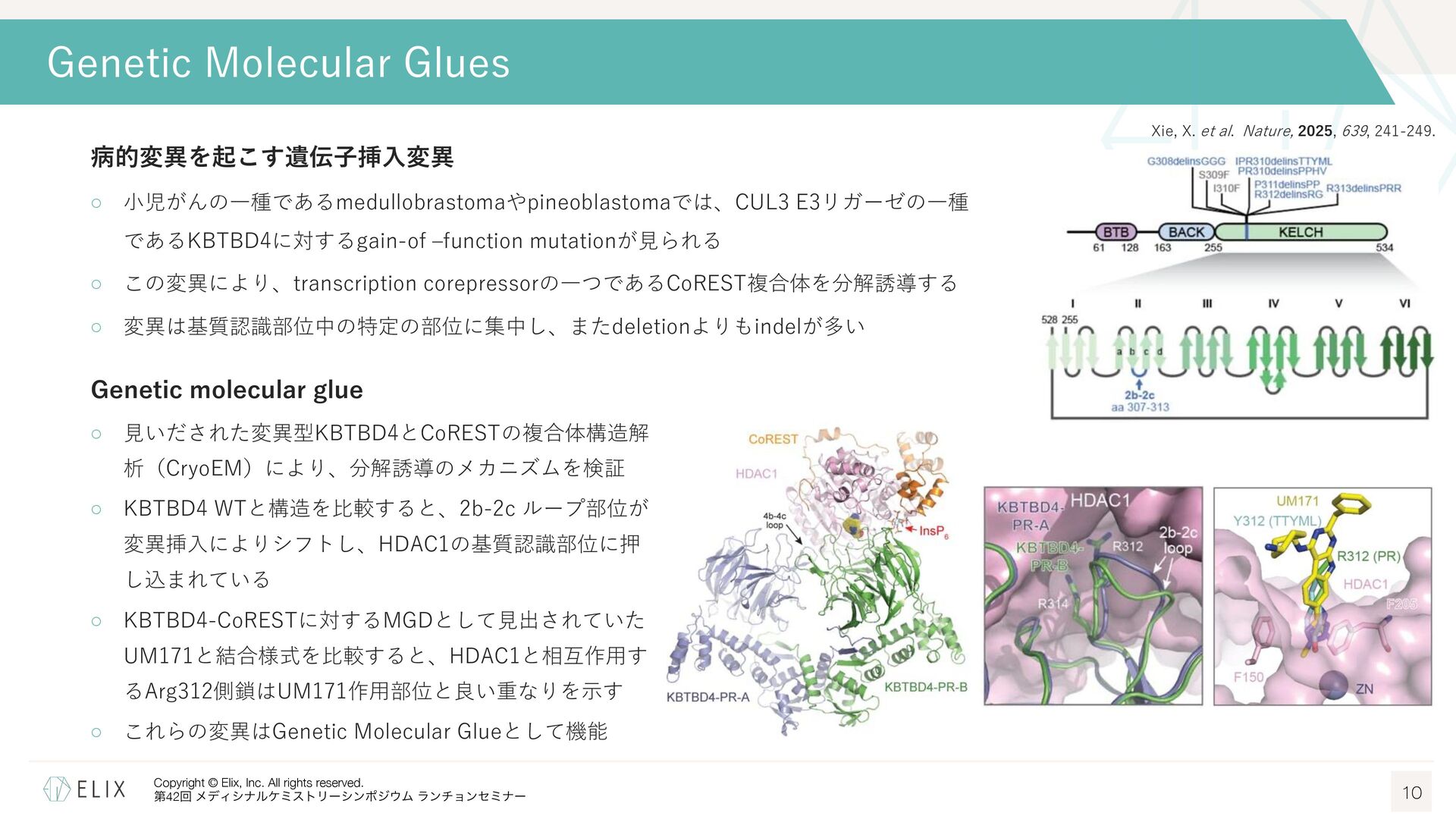{kind=link}
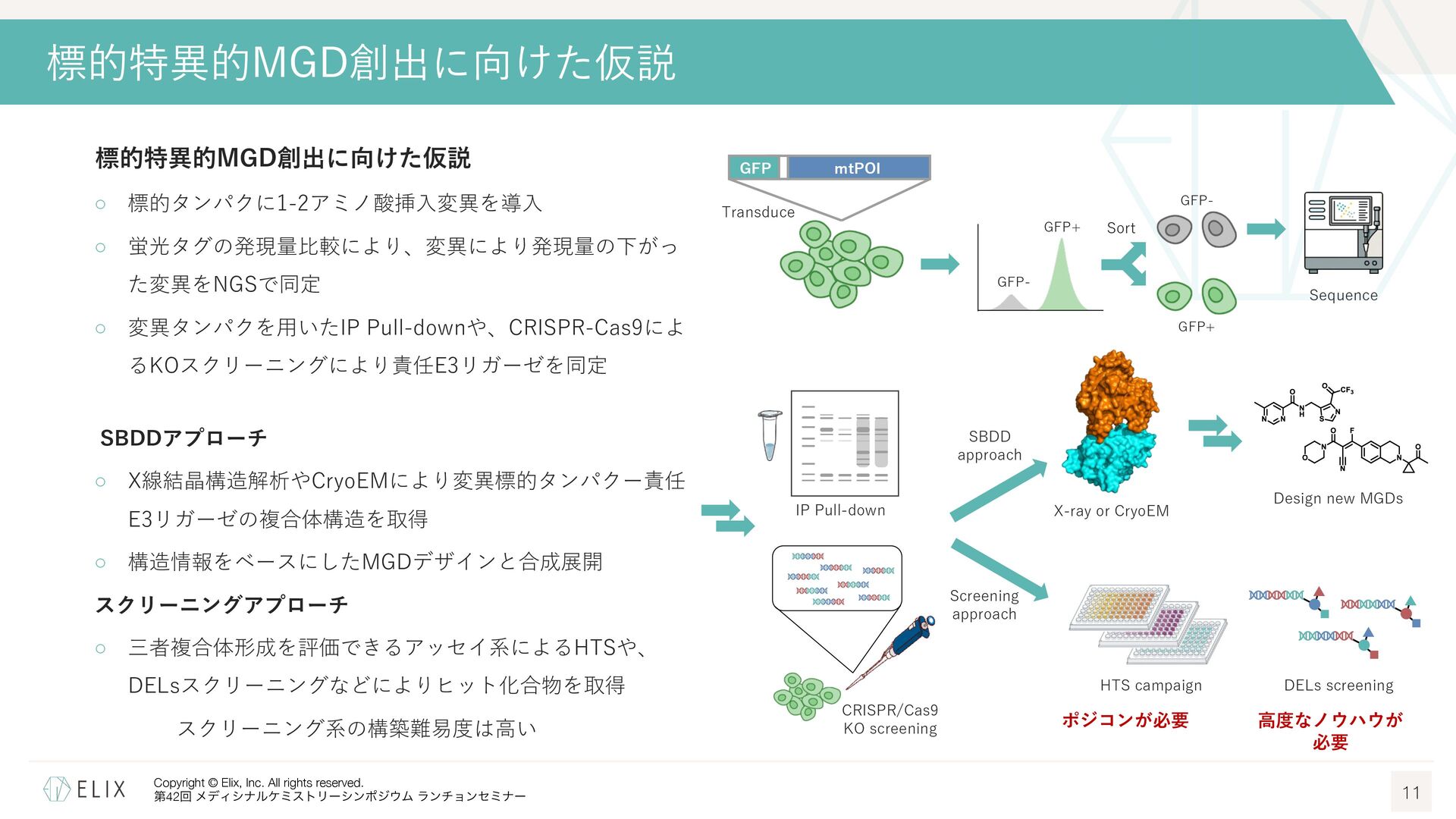{kind=link}
{kind=link}
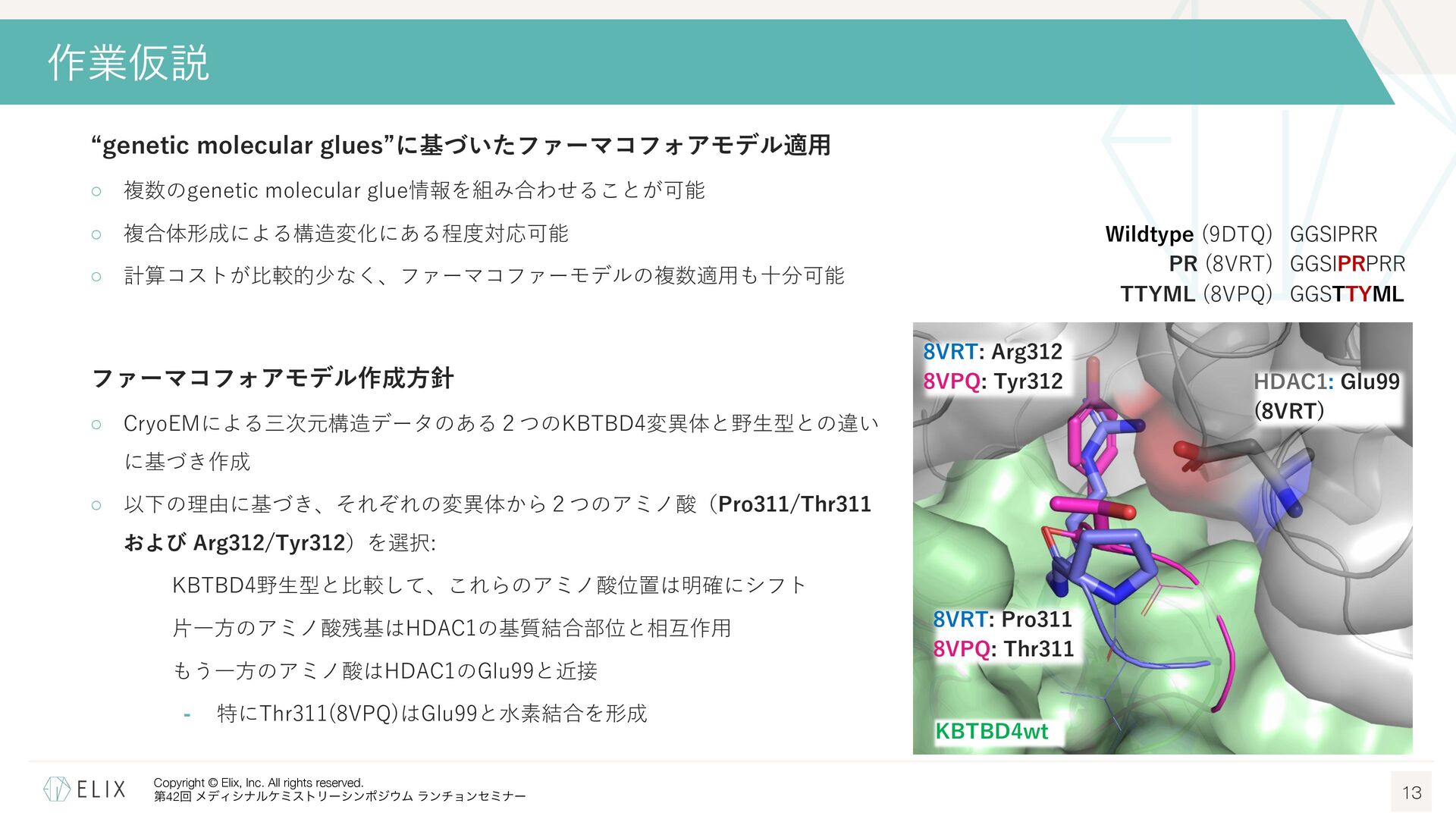{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}