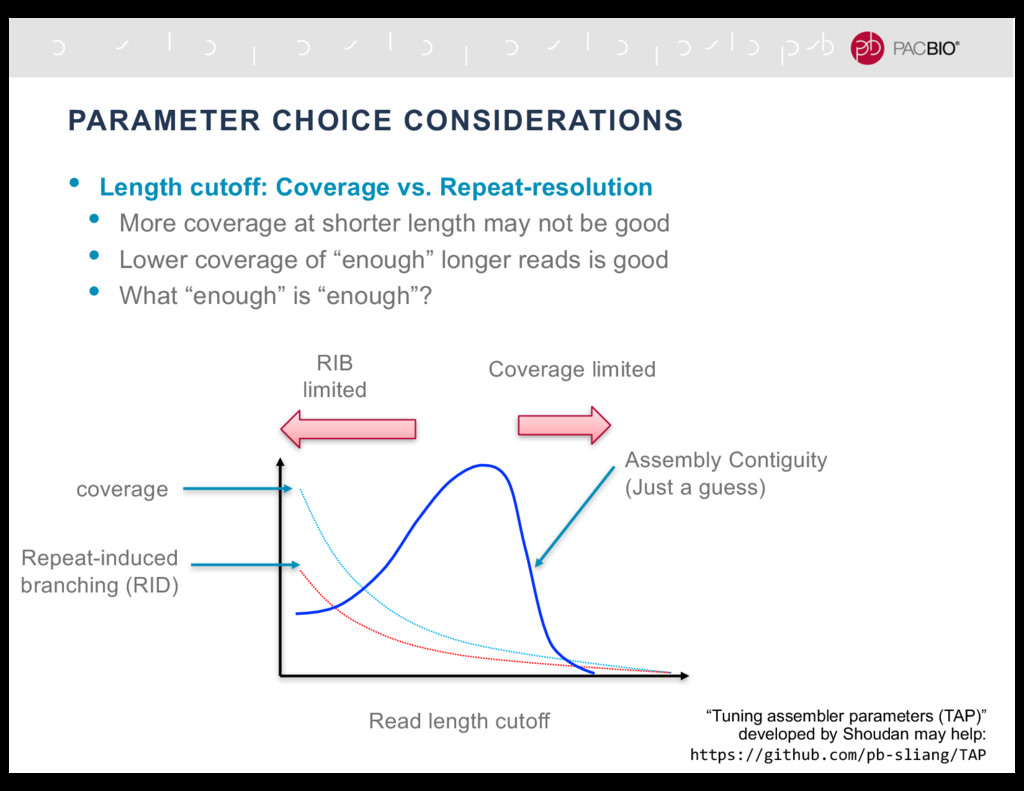
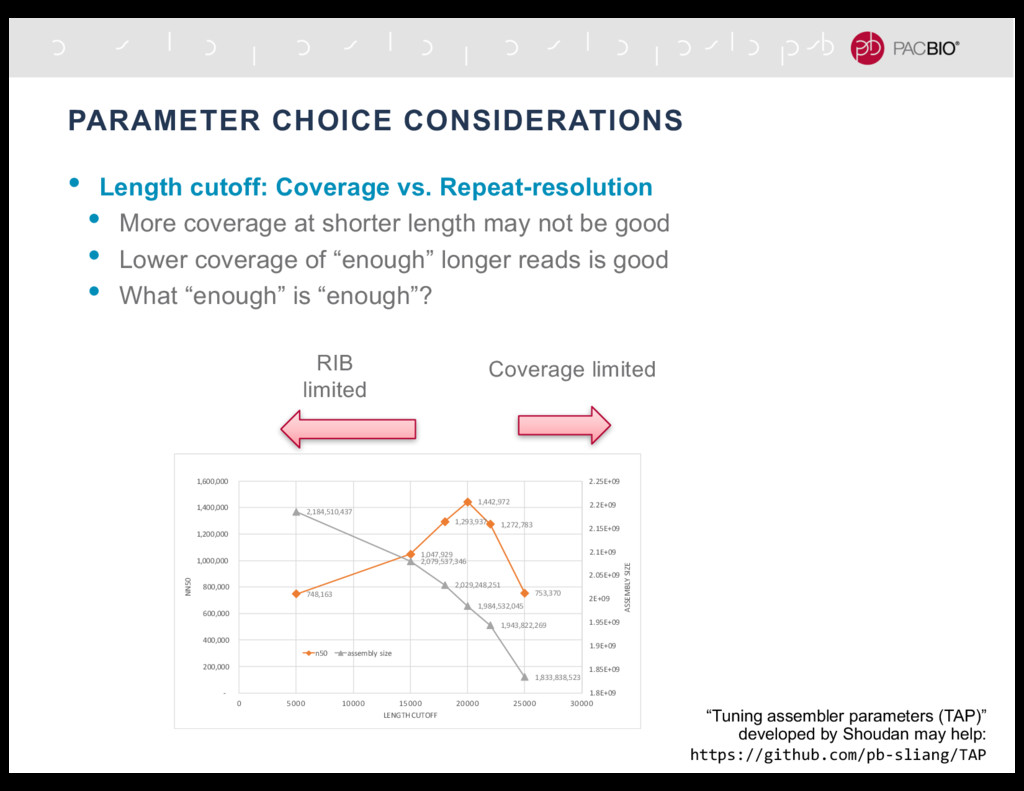
More coverage at shorter length may not be good • Lower coverage of “enough” longer reads is good • What “enough” is “enough”? RIB limited Coverage limited “Tuning assembler parameters (TAP)” developed by Shoudan may help: https://github.com/pb-sliang/TAP 748,163 1,047,929 1,293,937 1,442,972 1,272,783 753,370 2,184,510,437 2,079,537,346 2,029,248,251 1,984,532,045 1,943,822,269 1,833,838,523 1.8E+09 1.85E+09 1.9E+09 1.95E+09 2E+09 2.05E+09 2.1E+09 2.15E+09 2.2E+09 2.25E+09 - 200,000 400,000 600,000 800,000 1,000,000 1,200,000 1,400,000 1,600,000 0 5000 10000 15000 20000 25000 30000 ASSEMBLY SIZE NN50 LENGTH CUTOFF n50 assembly size
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}