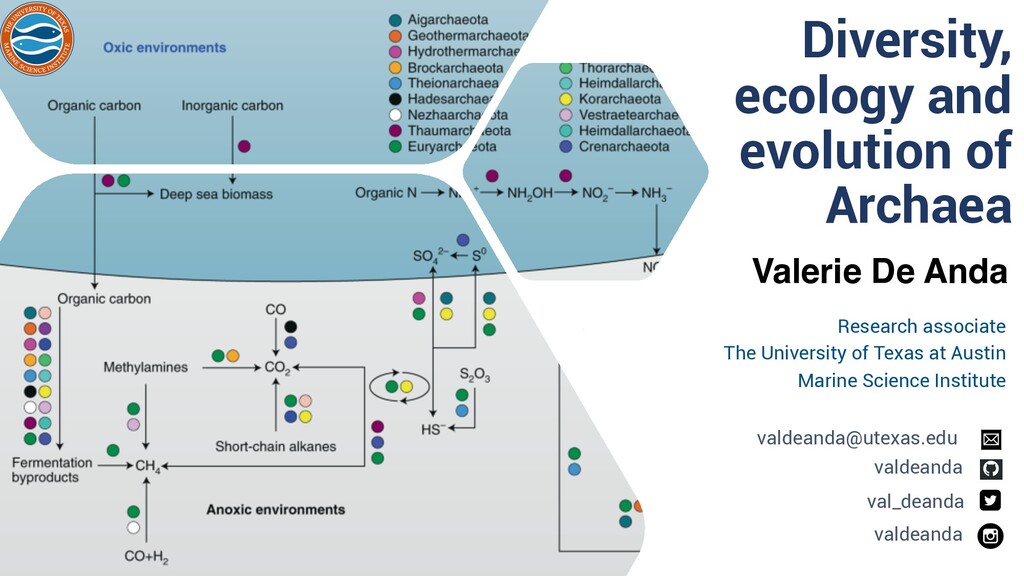
Invited seminar talk for the Laboratory of Francisco Bolivar Zapata and Adelfo Escalante and from the Biotechnology Institute Cell Engineering and Biocatalysis Department, National Autonomous University of Mexico.
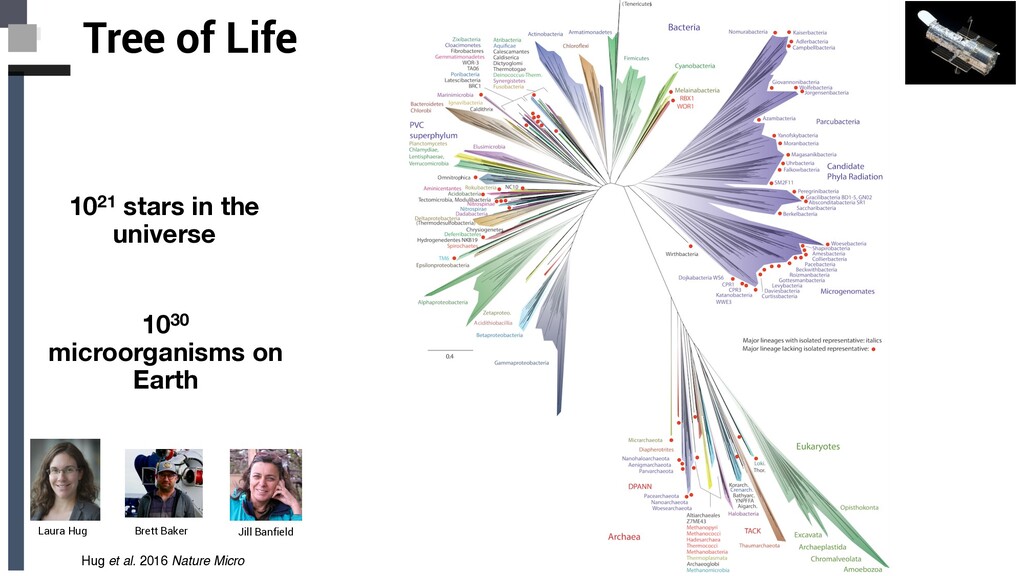
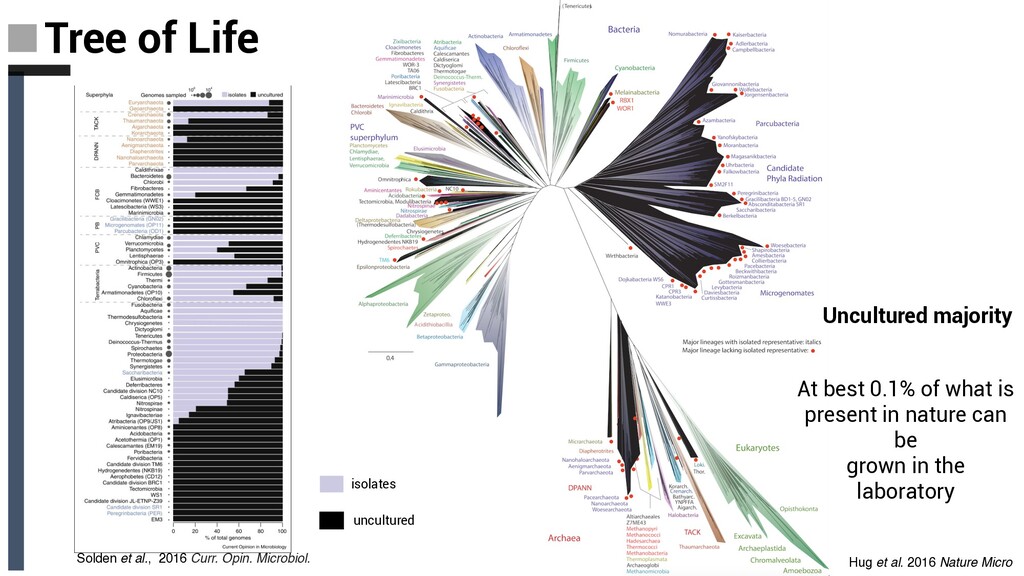
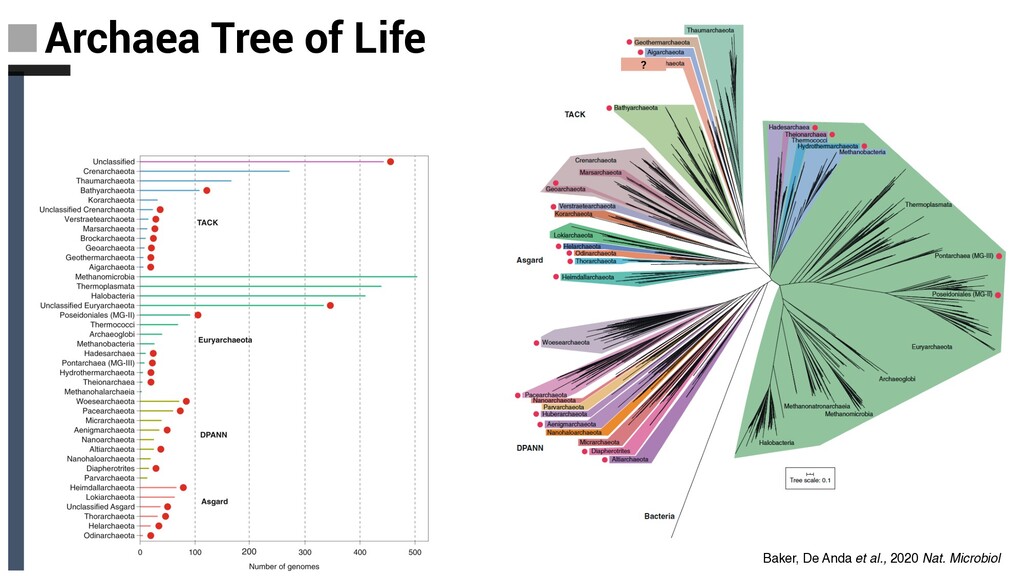
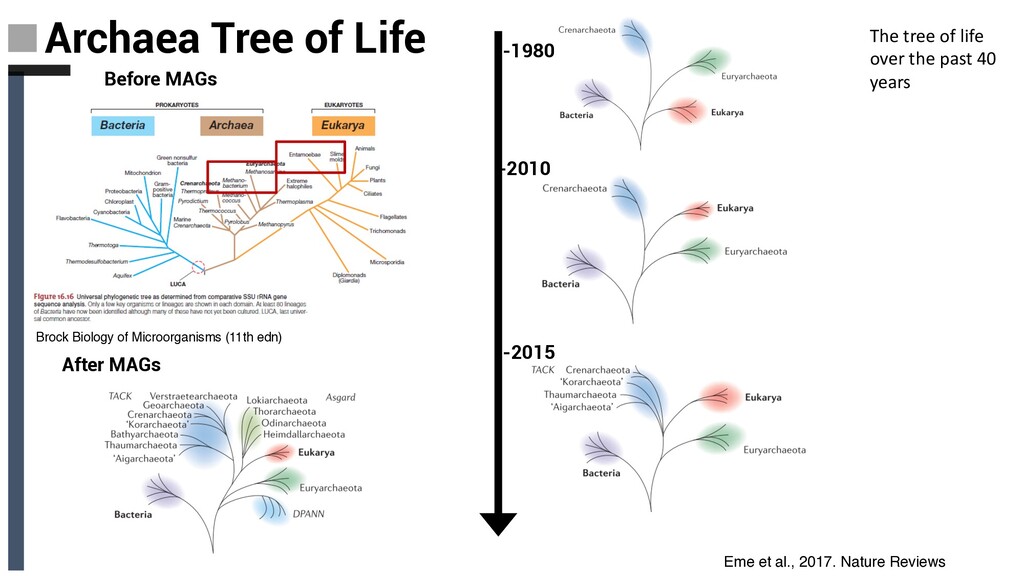
Tree of Life ❏ Archaeal tree of life Euryarchaea, DPANN,TACK and Asgard: § Discovery Novel Lineages § Ecology § Metabolism § Evolutionary implications ❏ Concluding Remarks and Future Perpectives Content
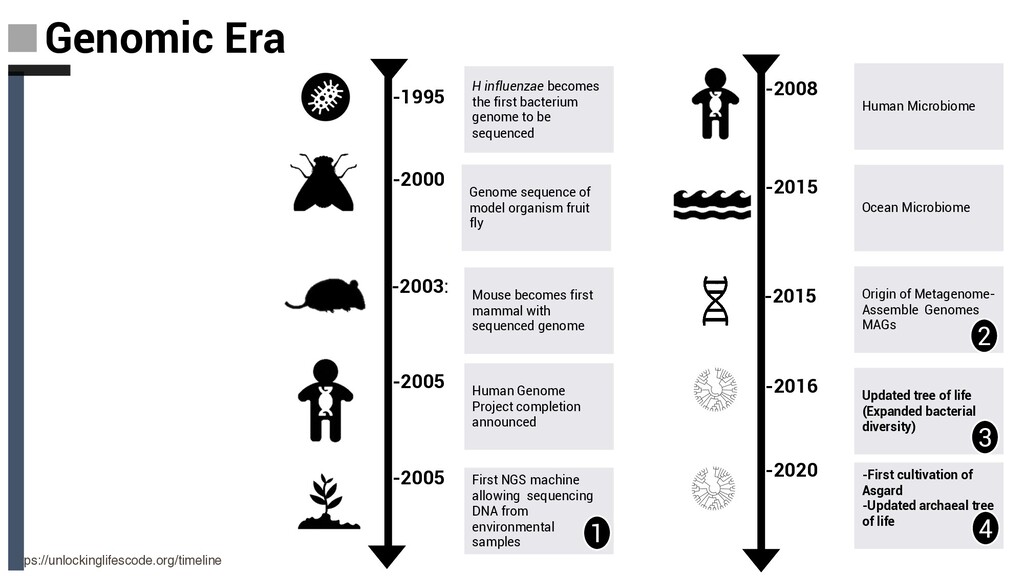
first mammal with sequenced genome Human Genome Project completion announced First NGS machine allowing sequencing DNA from environmental samples H influenzae becomes the first bacterium genome to be sequenced -1995 -2000 -2003: -2005 -2005 Human Microbiome Ocean Microbiome -2008 -2015 -2015 -2016 -2020 Origin of Metagenome- Assemble Genomes MAGs Updated tree of life (Expanded bacterial diversity) -First cultivation of Asgard -Updated archaeal tree of life 2 3 4 1 Genomic Era
Tree of Life ❏ Archaeal tree of life Euryarchaea, DPANN,TACK and Asgard: § Discovery Novel Lineages § Ecology § Metabolism § Evolutionary implications ❏ Concluding Remarks and Future Perpectives Content Diversity, ecology and evolution of Archaea
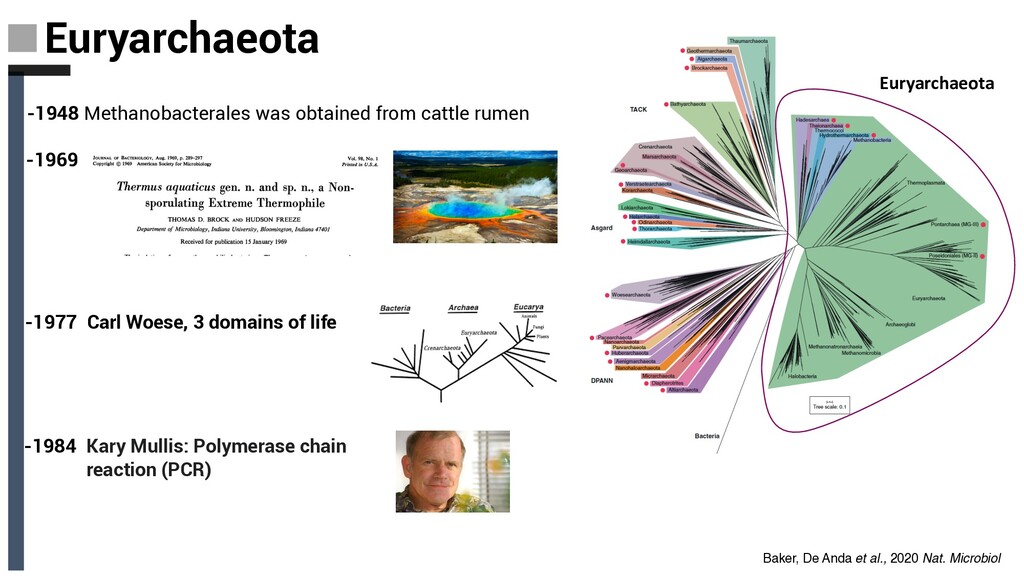
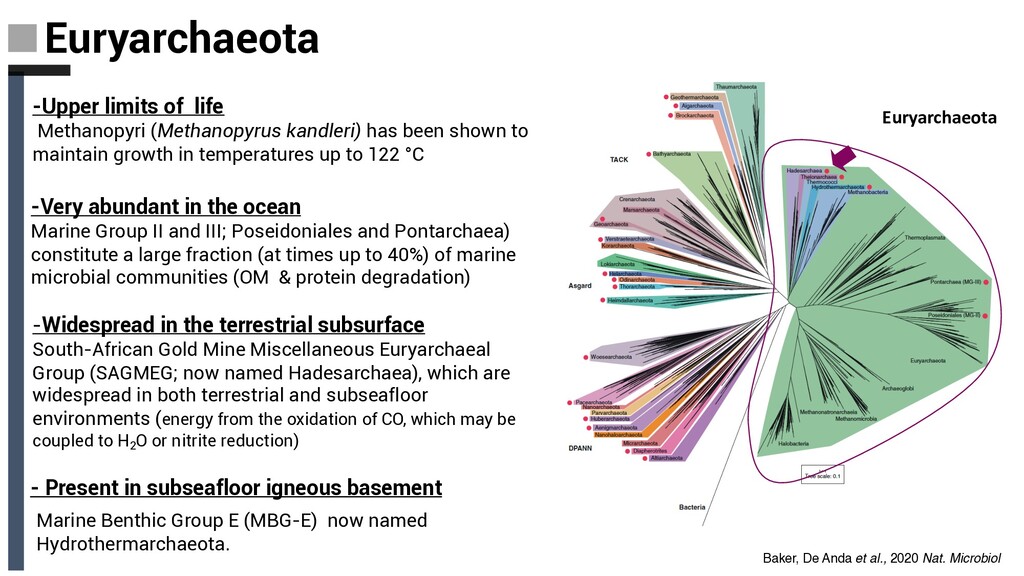
shown to maintain growth in temperatures up to 122 °C -Very abundant in the ocean Marine Group II and III; Poseidoniales and Pontarchaea) constitute a large fraction (at times up to 40%) of marine microbial communities (OM & protein degradation) -Widespread in the terrestrial subsurface South-African Gold Mine Miscellaneous Euryarchaeal Group (SAGMEG; now named Hadesarchaea), which are widespread in both terrestrial and subseafloor environments (energy from the oxidation of CO, which may be coupled to H2 O or nitrite reduction) - Present in subseafloor igneous basement Marine Benthic Group E (MBG-E) now named Hydrothermarchaeota. Euryarchaeota Baker, De Anda et al., 2020 Nat. Microbiol
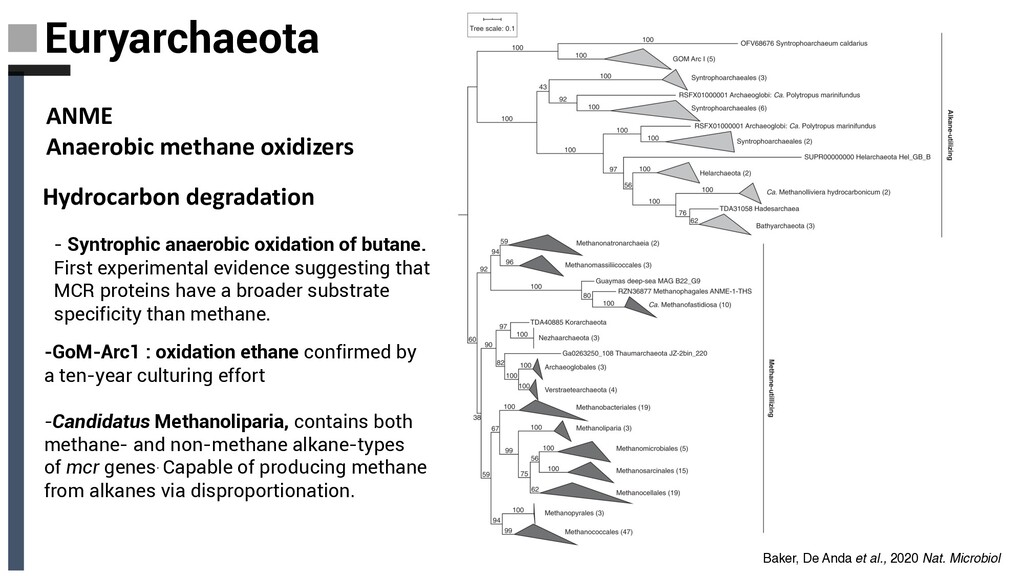
experimental evidence suggesting that MCR proteins have a broader substrate specificity than methane. -GoM-Arc1 : oxidation ethane confirmed by a ten-year culturing effort -Candidatus Methanoliparia, contains both methane- and non-methane alkane-types of mcr genes. Capable of producing methane from alkanes via disproportionation. Baker, De Anda et al., 2020 Nat. Microbiol ANME Anaerobic methane oxidizers
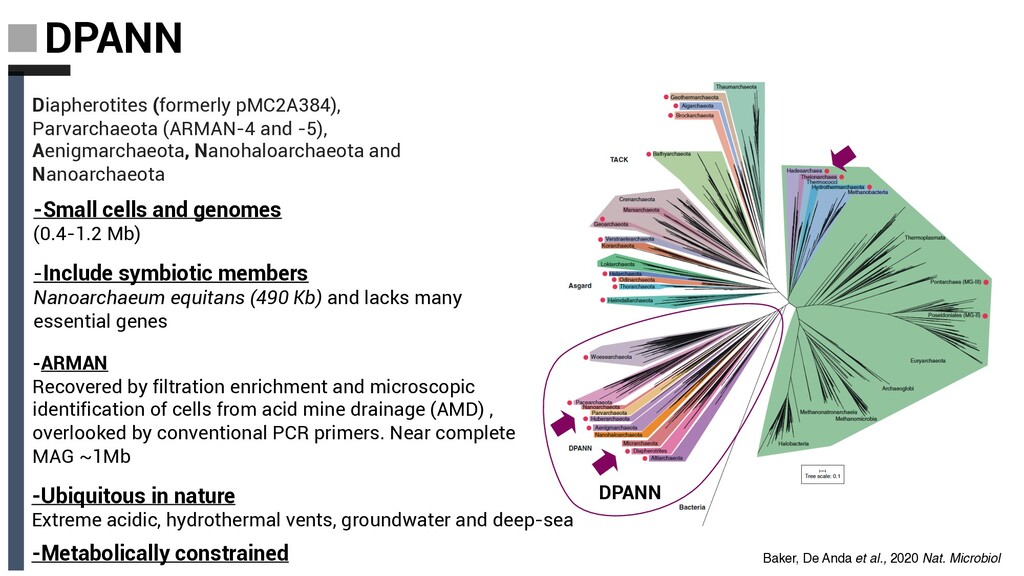
and Nanoarchaeota -Small cells and genomes (0.4-1.2 Mb) -ARMAN Recovered by filtration enrichment and microscopic identification of cells from acid mine drainage (AMD) , overlooked by conventional PCR primers. Near complete MAG ~1Mb -Include symbiotic members Nanoarchaeum equitans (490 Kb) and lacks many essential genes -Ubiquitous in nature Extreme acidic, hydrothermal vents, groundwater and deep-sea -Metabolically constrained DPANN Baker, De Anda et al., 2020 Nat. Microbiol
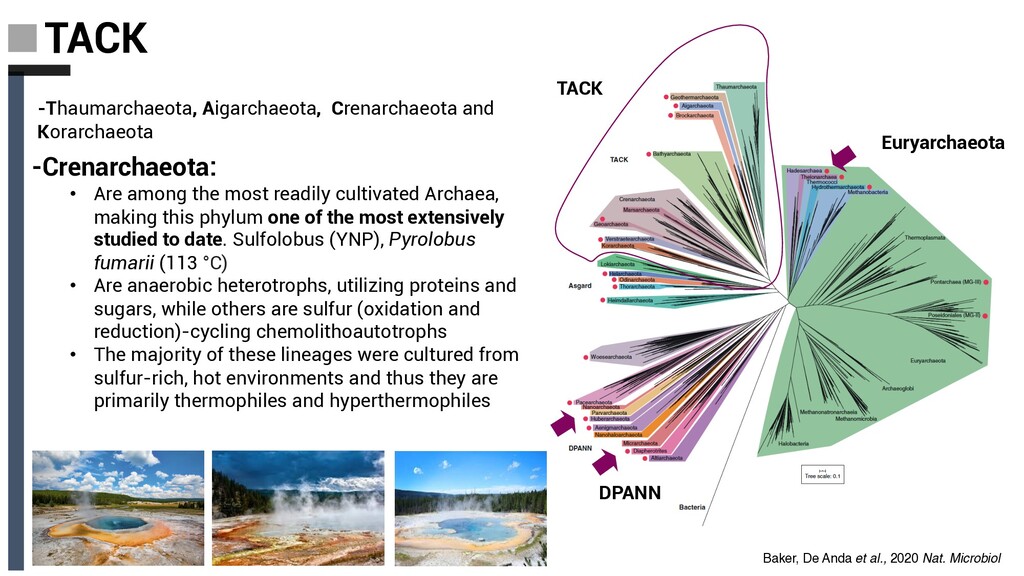
making this phylum one of the most extensively studied to date. Sulfolobus (YNP), Pyrolobus fumarii (113 °C) -Thaumarchaeota, Aigarchaeota, Crenarchaeota and Korarchaeota DPANN Euryarchaeota • Are anaerobic heterotrophs, utilizing proteins and sugars, while others are sulfur (oxidation and reduction)-cycling chemolithoautotrophs • The majority of these lineages were cultured from sulfur-rich, hot environments and thus they are primarily thermophiles and hyperthermophiles TACK Baker, De Anda et al., 2020 Nat. Microbiol
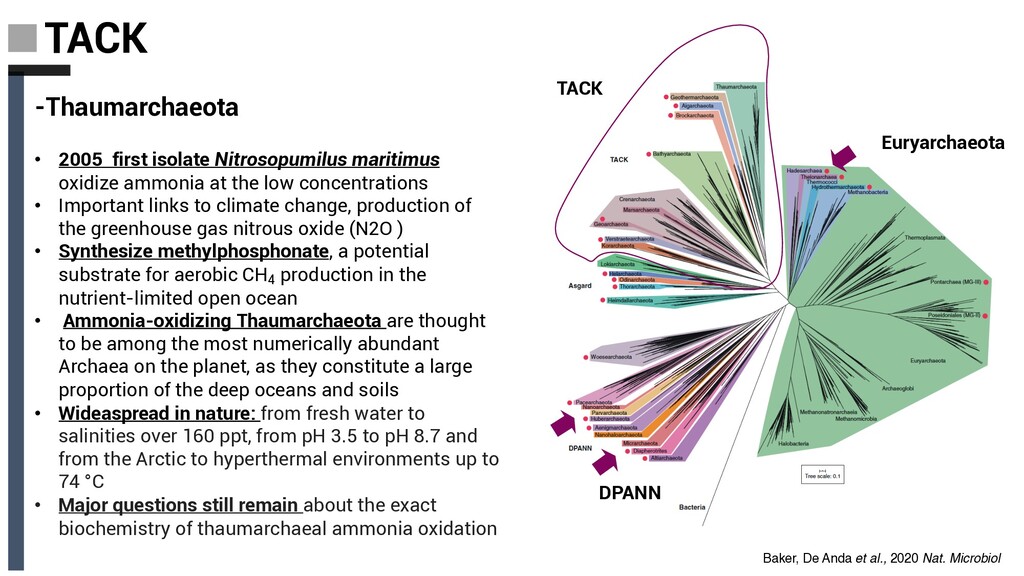
maritimus oxidize ammonia at the low concentrations • Important links to climate change, production of the greenhouse gas nitrous oxide (N2O ) • Synthesize methylphosphonate, a potential substrate for aerobic CH4 production in the nutrient-limited open ocean • Ammonia-oxidizing Thaumarchaeota are thought to be among the most numerically abundant Archaea on the planet, as they constitute a large proportion of the deep oceans and soils • Wideaspread in nature: from fresh water to salinities over 160 ppt, from pH 3.5 to pH 8.7 and from the Arctic to hyperthermal environments up to 74 °C • Major questions still remain about the exact biochemistry of thaumarchaeal ammonia oxidation Baker, De Anda et al., 2020 Nat. Microbiol
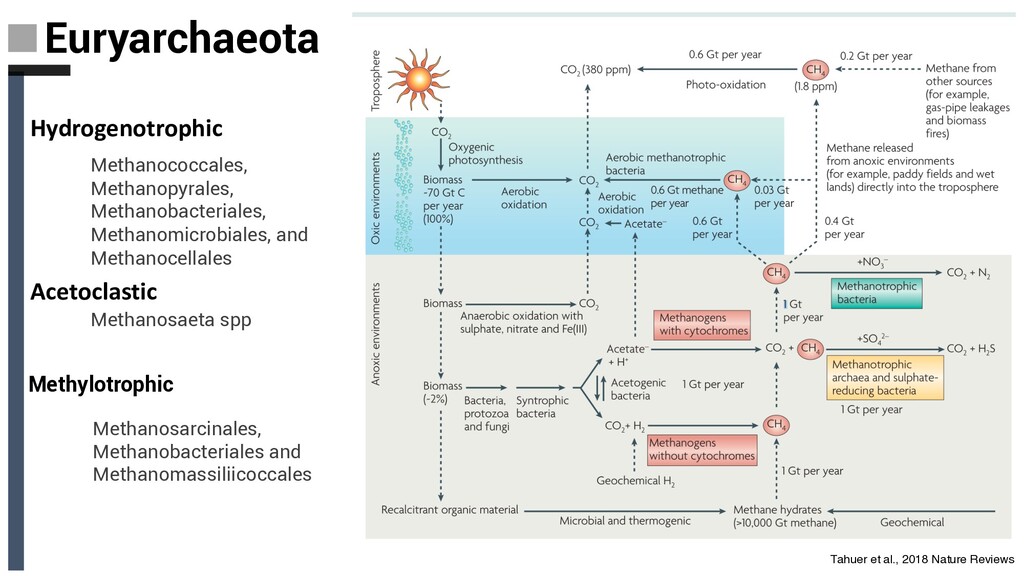
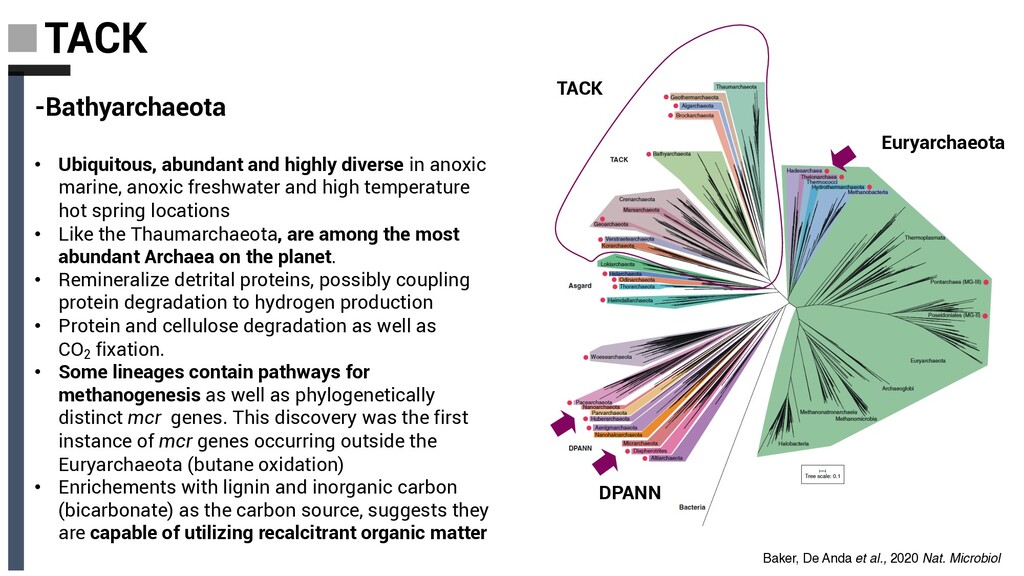
diverse in anoxic marine, anoxic freshwater and high temperature hot spring locations • Like the Thaumarchaeota, are among the most abundant Archaea on the planet. • Remineralize detrital proteins, possibly coupling protein degradation to hydrogen production • Protein and cellulose degradation as well as CO2 fixation. • Some lineages contain pathways for methanogenesis as well as phylogenetically distinct mcr genes. This discovery was the first instance of mcr genes occurring outside the Euryarchaeota (butane oxidation) • Enrichements with lignin and inorganic carbon (bicarbonate) as the carbon source, suggests they are capable of utilizing recalcitrant organic matter Baker, De Anda et al., 2020 Nat. Microbiol
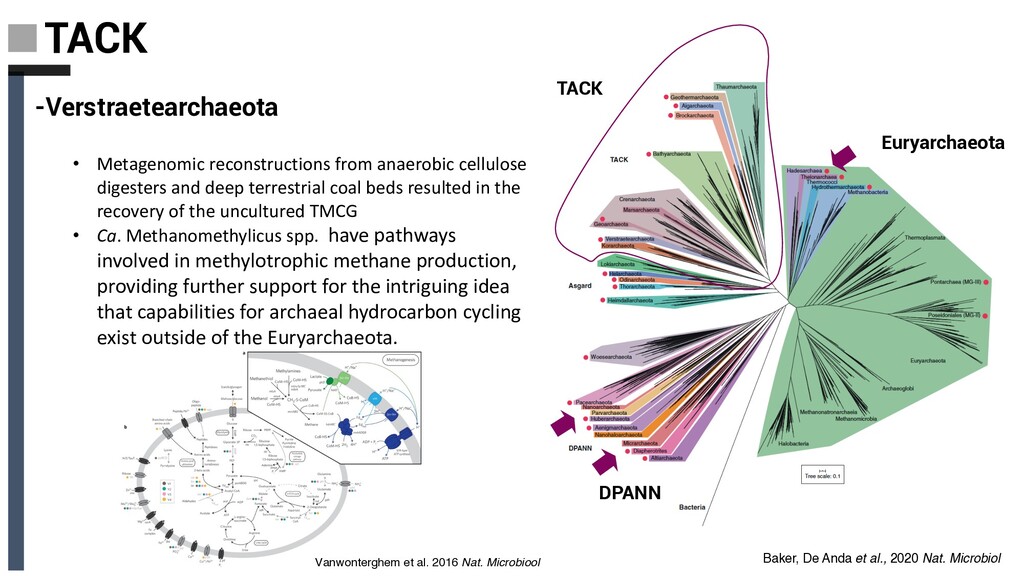
cellulose digesters and deep terrestrial coal beds resulted in the recovery of the uncultured TMCG • Ca. Methanomethylicus spp. have pathways involved in methylotrophic methane production, providing further support for the intriguing idea that capabilities for archaeal hydrocarbon cycling exist outside of the Euryarchaeota. Vanwonterghem et al. 2016 Nat. Microbiool Baker, De Anda et al., 2020 Nat. Microbiol
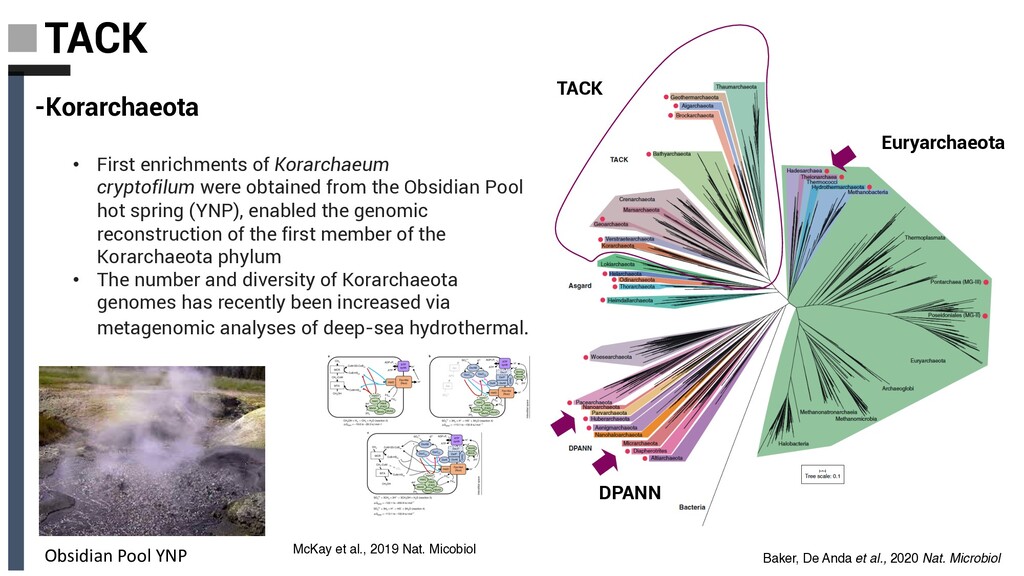
cryptofilum were obtained from the Obsidian Pool hot spring (YNP), enabled the genomic reconstruction of the first member of the Korarchaeota phylum • The number and diversity of Korarchaeota genomes has recently been increased via metagenomic analyses of deep-sea hydrothermal. Obsidian Pool YNP McKay et al., 2019 Nat. Micobiol Baker, De Anda et al., 2020 Nat. Microbiol
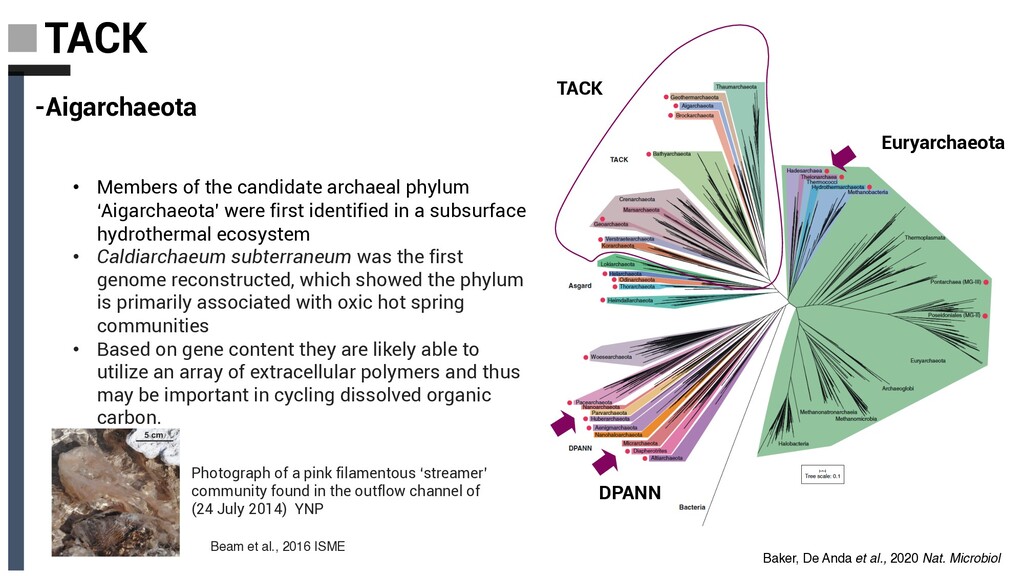
archaeal phylum ‘Aigarchaeota’ were first identified in a subsurface hydrothermal ecosystem • Caldiarchaeum subterraneum was the first genome reconstructed, which showed the phylum is primarily associated with oxic hot spring communities • Based on gene content they are likely able to utilize an array of extracellular polymers and thus may be important in cycling dissolved organic carbon. Photograph of a pink filamentous ‘streamer’ community found in the outflow channel of (24 July 2014) YNP Beam et al., 2016 ISME Baker, De Anda et al., 2020 Nat. Microbiol
in geothermal iron oxide microbial mats • Microaerobic geothermal Fe(III) oxide microbial mats across a temperature range from ~50–80 °C • Chemoorganotrophs and utilize a variety of reduced carbon substrates. Sulfonate may serve as a carbon source via a F420 - dependent butanal metabolism, representing s an additional mechanism for generating acetyl-CoA in archaea. Jay et al., Nat. Microbiol Baker, De Anda et al., 2020 Nat. Microbiol
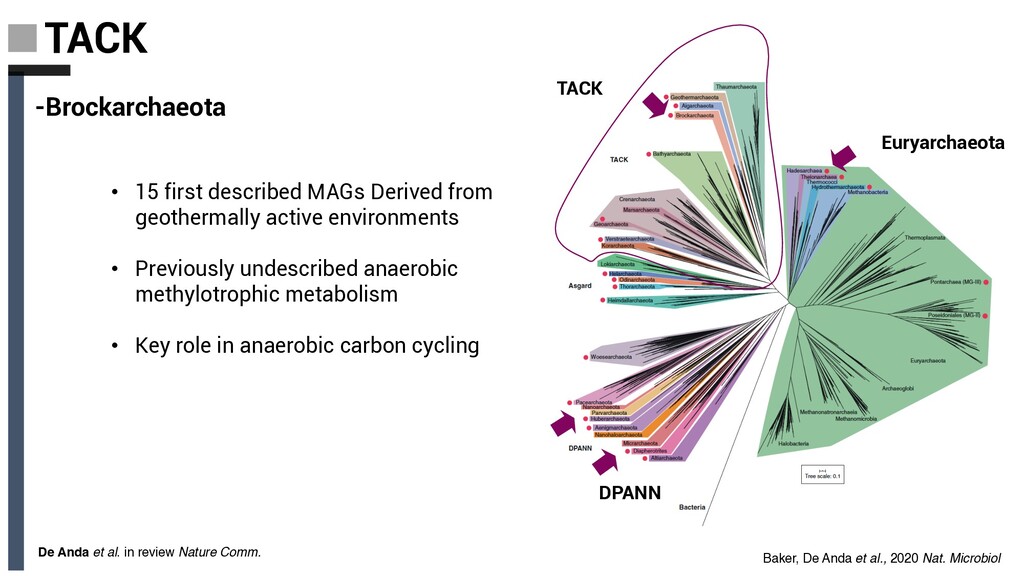
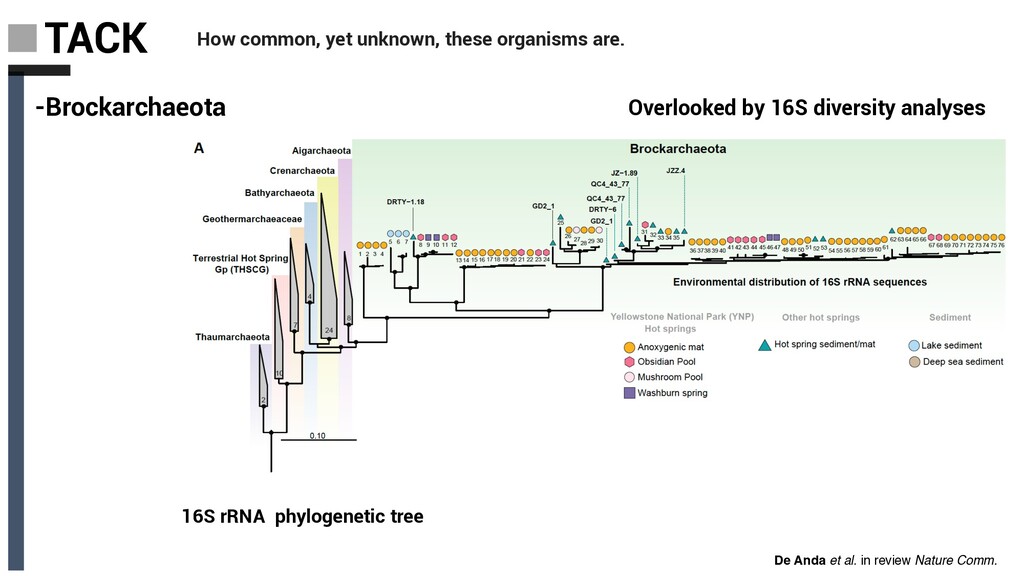
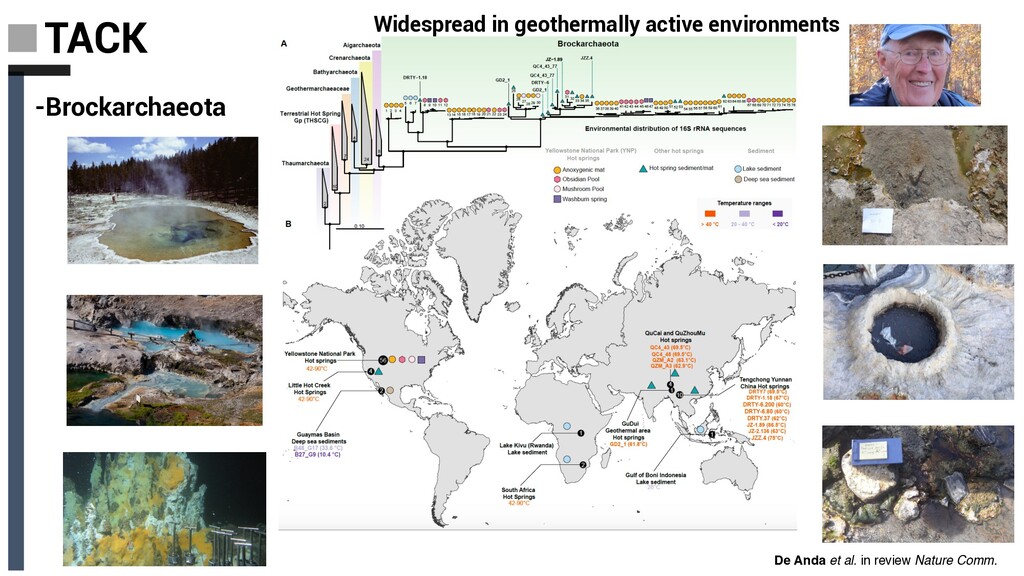
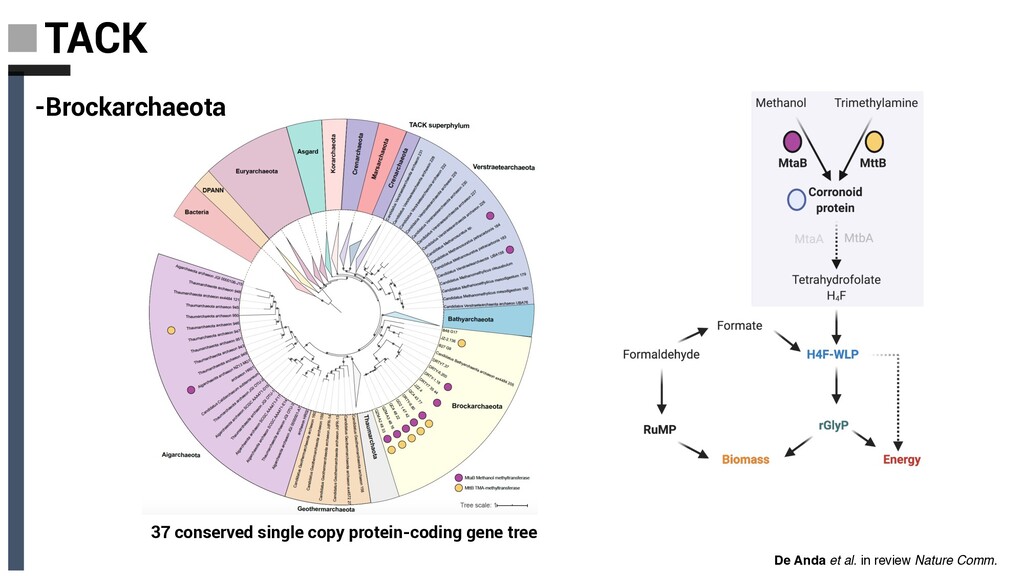
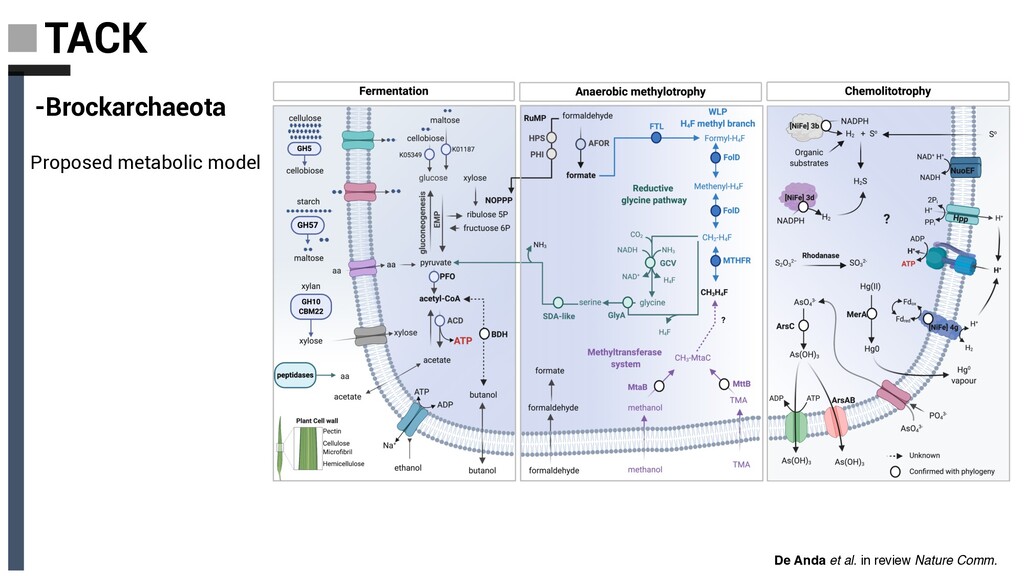
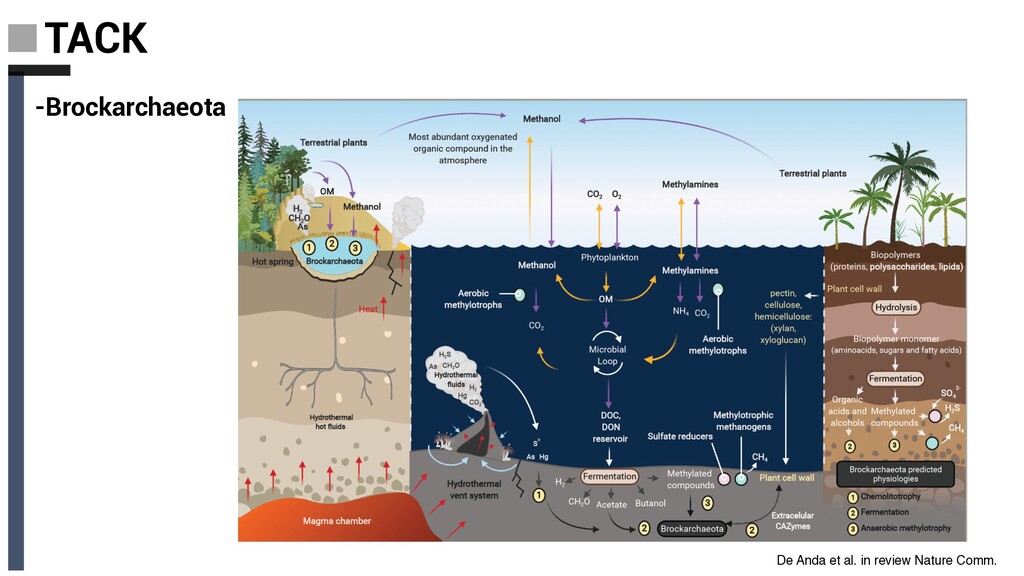
Derived from geothermally active environments • Previously undescribed anaerobic methylotrophic metabolism • Key role in anaerobic carbon cycling De Anda et al. in review Nature Comm. Baker, De Anda et al., 2020 Nat. Microbiol
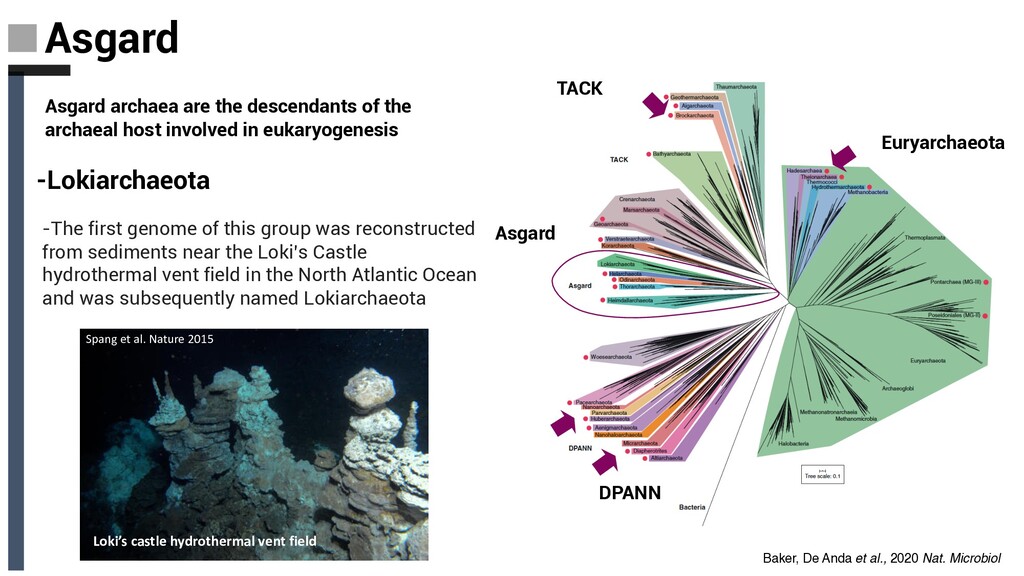
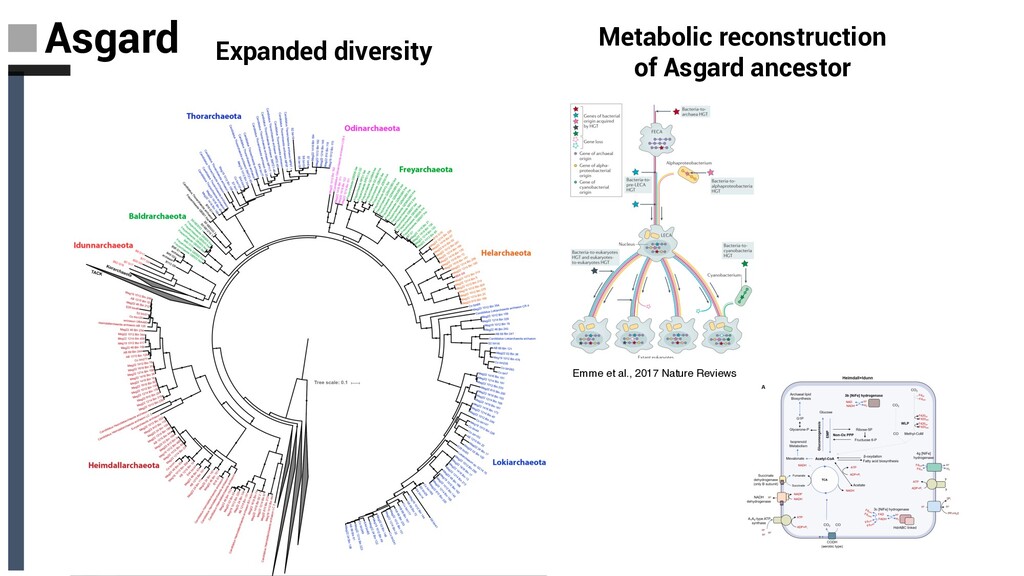
Spang et al. Nature 2015 -The first genome of this group was reconstructed from sediments near the Loki’s Castle hydrothermal vent field in the North Atlantic Ocean and was subsequently named Lokiarchaeota -Lokiarchaeota Asgard archaea are the descendants of the archaeal host involved in eukaryogenesis Baker, De Anda et al., 2020 Nat. Microbiol
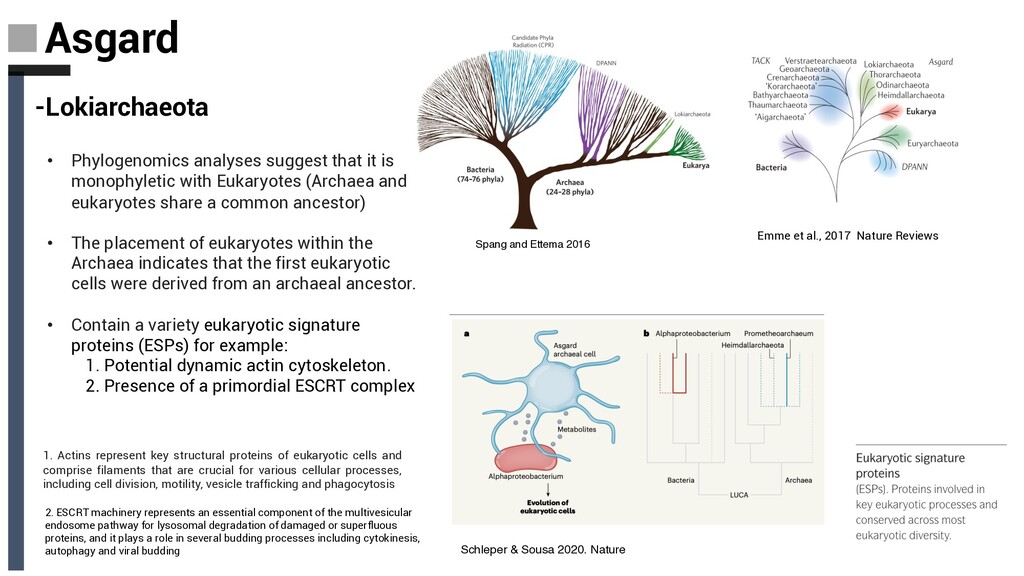
with Eukaryotes (Archaea and eukaryotes share a common ancestor) • The placement of eukaryotes within the Archaea indicates that the first eukaryotic cells were derived from an archaeal ancestor. • Contain a variety eukaryotic signature proteins (ESPs) for example: 1. Potential dynamic actin cytoskeleton. 2. Presence of a primordial ESCRT complex Schleper & Sousa 2020. Nature 1. Actins represent key structural proteins of eukaryotic cells and comprise filaments that are crucial for various cellular processes, including cell division, motility, vesicle trafficking and phagocytosis 2. ESCRT machinery represents an essential component of the multivesicular endosome pathway for lysosomal degradation of damaged or superfluous proteins, and it plays a role in several budding processes including cytokinesis, autophagy and viral budding Emme et al., 2017 Nature Reviews Spang and Ettema 2016
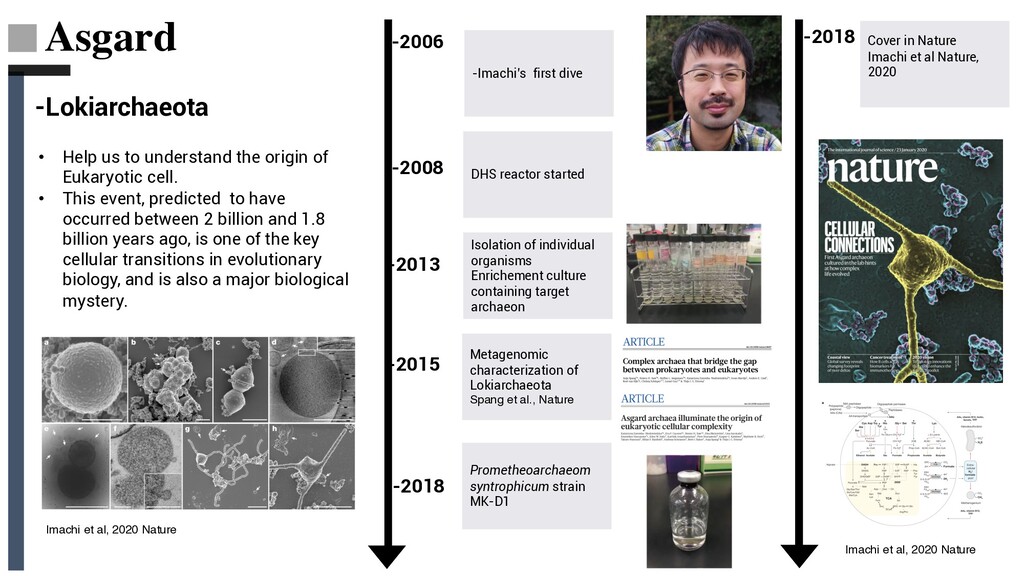
culture containing target archaeon Metagenomic characterization of Lokiarchaeota Spang et al., Nature Prometheoarchaeom syntrophicum strain MK-D1 -Imachi’s first dive -2006 -2008 -2013 -2015 -2018 Cover in Nature Imachi et al Nature, 2020 -2018 • Help us to understand the origin of Eukaryotic cell. • This event, predicted to have occurred between 2 billion and 1.8 billion years ago, is one of the key cellular transitions in evolutionary biology, and is also a major biological mystery. Imachi et al, 2020 Nature Imachi et al, 2020 Nature
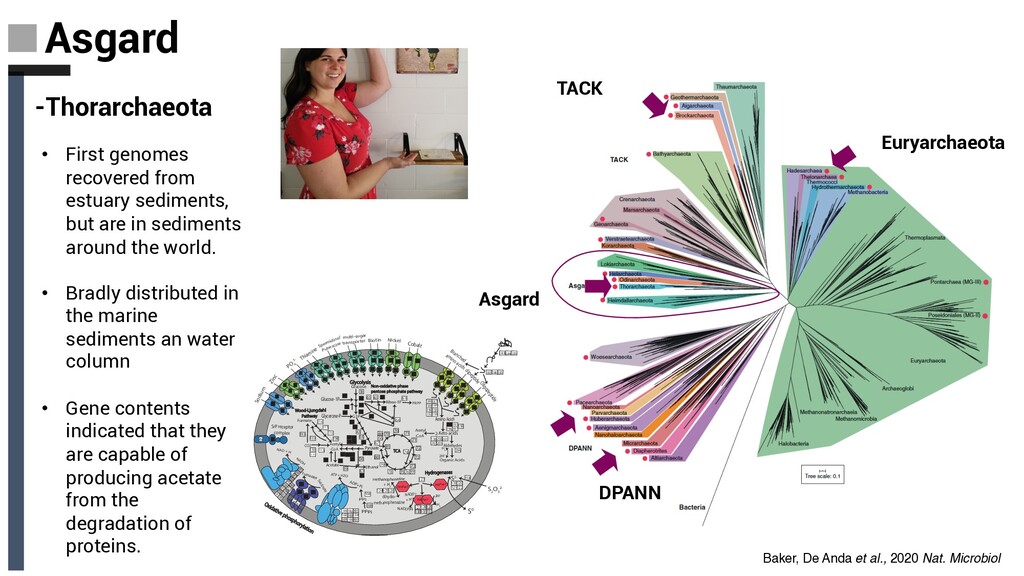
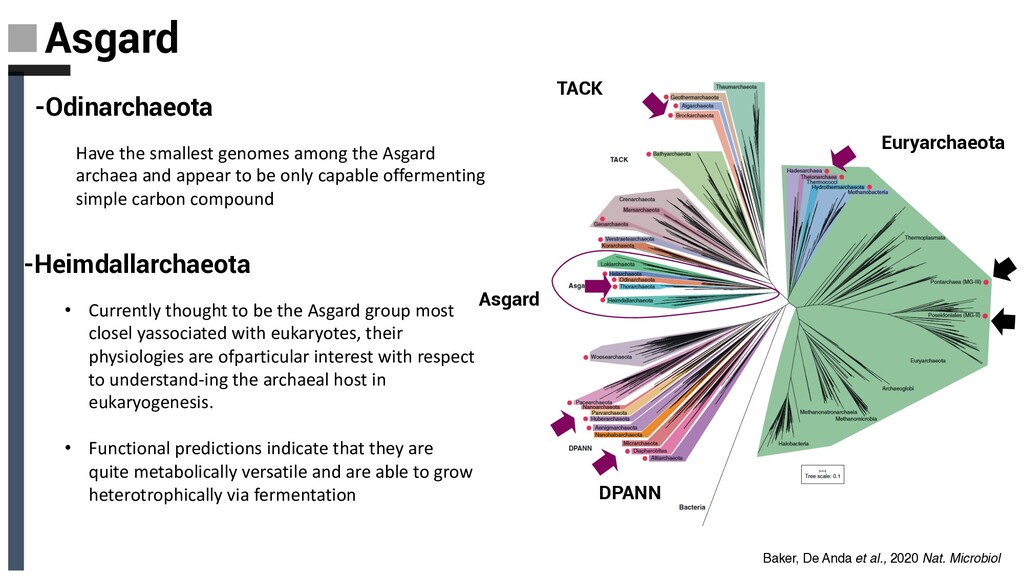
-Odinarchaeota Have the smallest genomes among the Asgard archaea and appear to be only capable offermenting simple carbon compound -Heimdallarchaeota • Currently thought to be the Asgard group most closel yassociated with eukaryotes, their physiologies are ofparticular interest with respect to understand-ing the archaeal host in eukaryogenesis. • Functional predictions indicate that they are quite metabolically versatile and are able to grow heterotrophically via fermentation Baker, De Anda et al., 2020 Nat. Microbiol
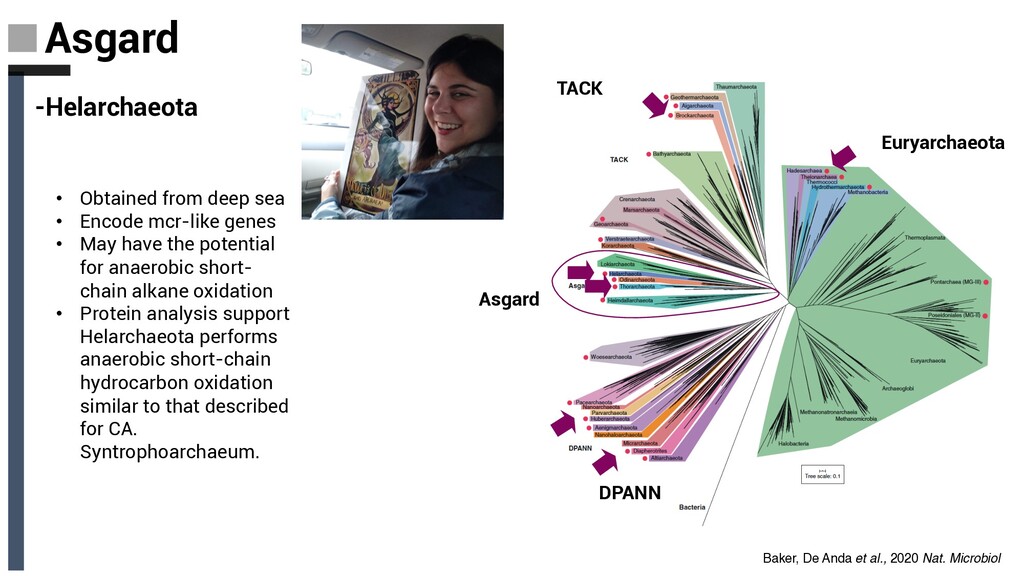
sea • Encode mcr-like genes • May have the potential for anaerobic short- chain alkane oxidation • Protein analysis support Helarchaeota performs anaerobic short-chain hydrocarbon oxidation similar to that described for CA. Syntrophoarchaeum. Baker, De Anda et al., 2020 Nat. Microbiol
of life Euryarchaea, DPANN,TACK and Asgard: § Discovery Novel Lineages § Ecology § Metabolism § Evolutionary implications ❏ Concluding Remarks and Future Perpectives Content Diversity, ecology and evolution of Archaea
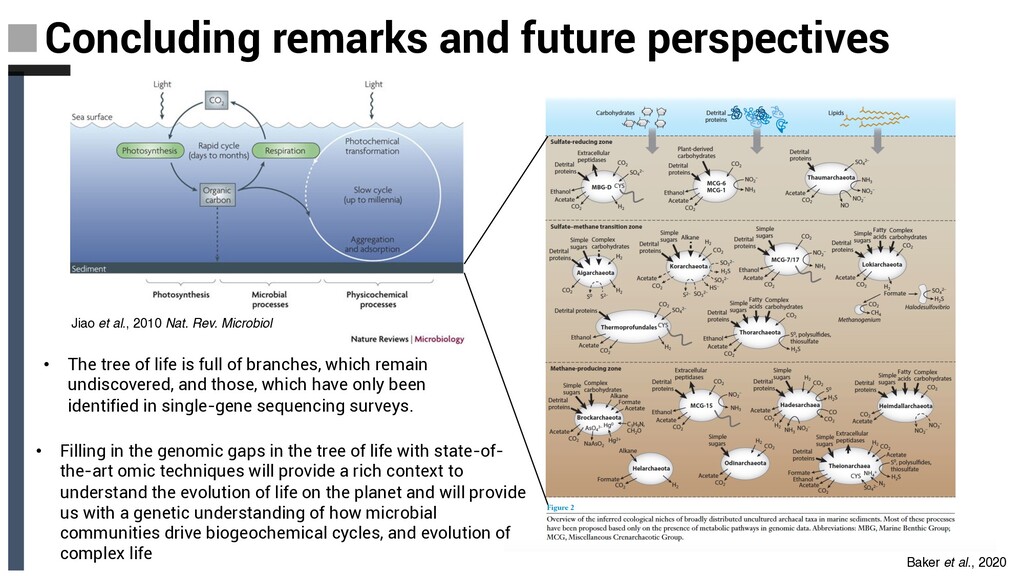
Microbiol Concluding remarks and future perspectives • The tree of life is full of branches, which remain undiscovered, and those, which have only been identified in single-gene sequencing surveys. • Filling in the genomic gaps in the tree of life with state-of- the-art omic techniques will provide a rich context to understand the evolution of life on the planet and will provide us with a genetic understanding of how microbial communities drive biogeochemical cycles, and evolution of complex life
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}