This is the slide deck of a talk I gave in #SFAF2015 (http://www.lanl.gov/conferences/sequencing-finishing-analysis-future/index.php) .
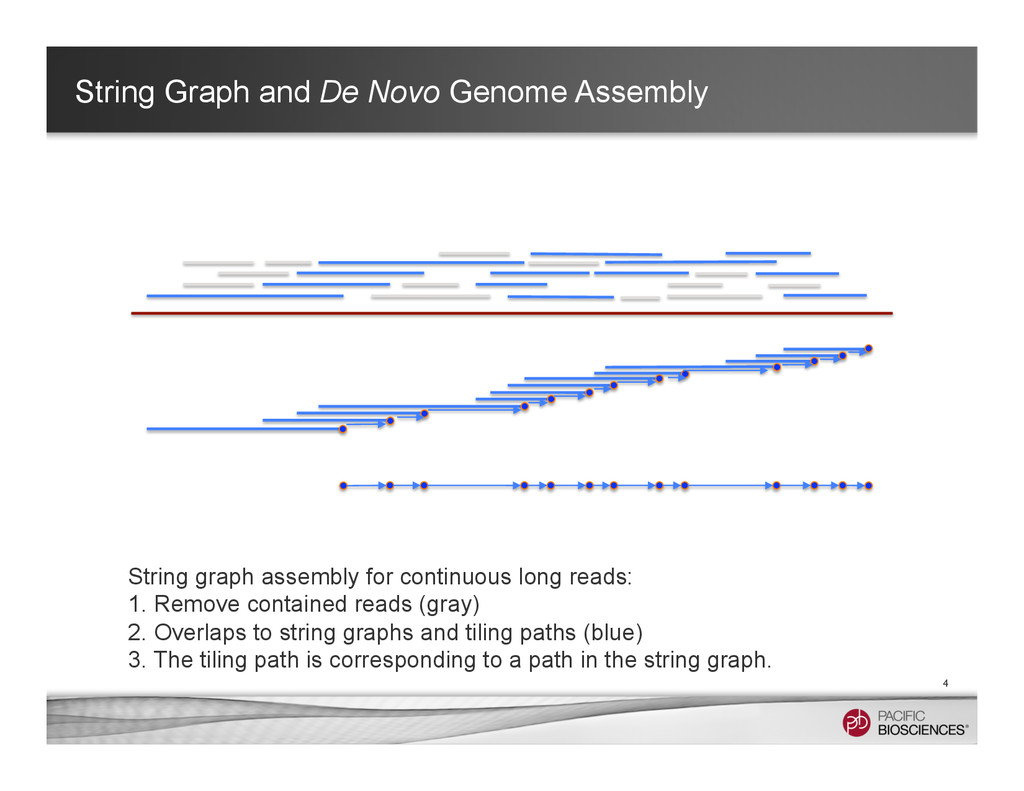
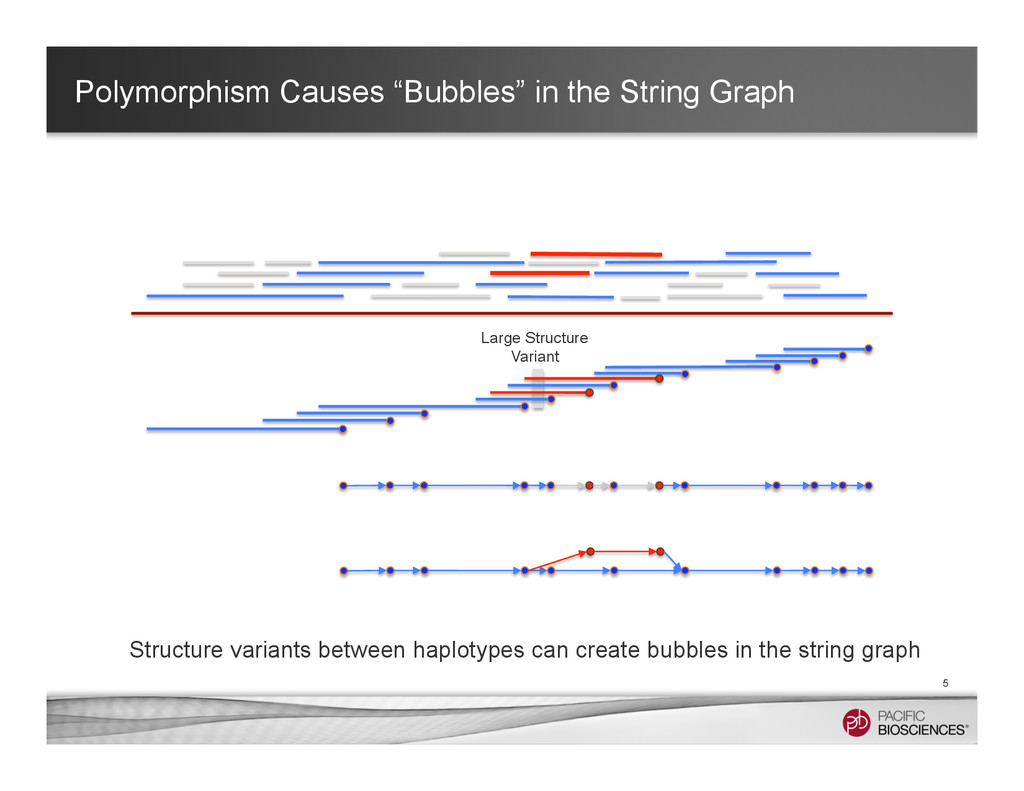
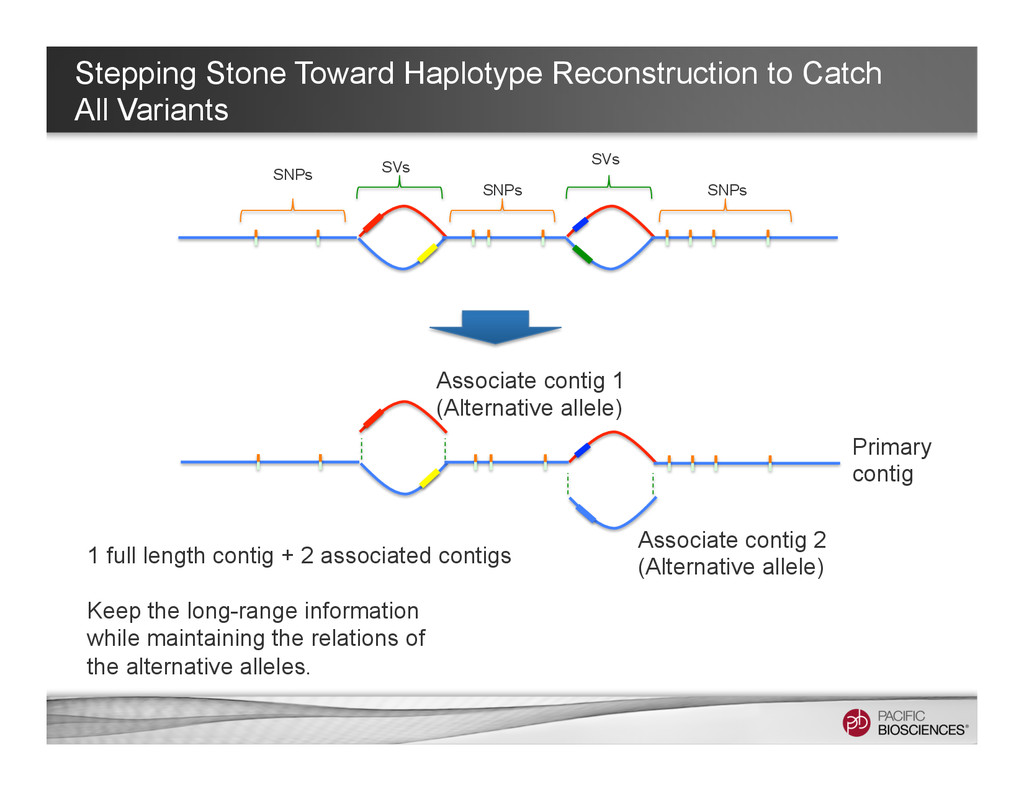
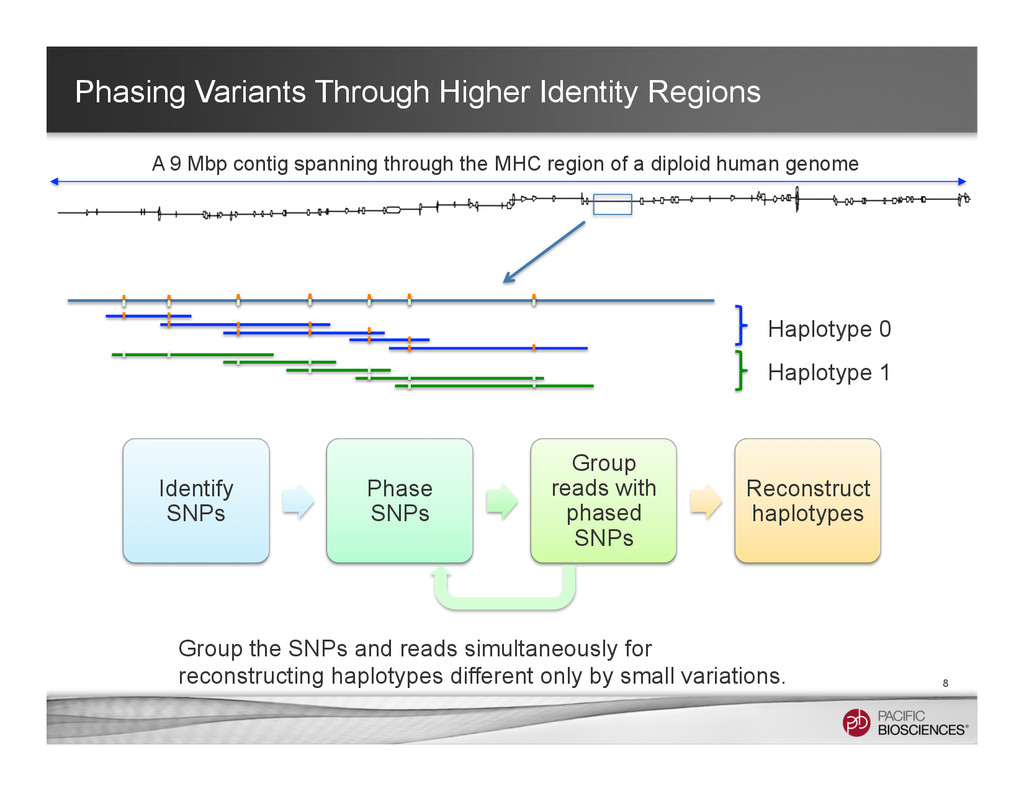
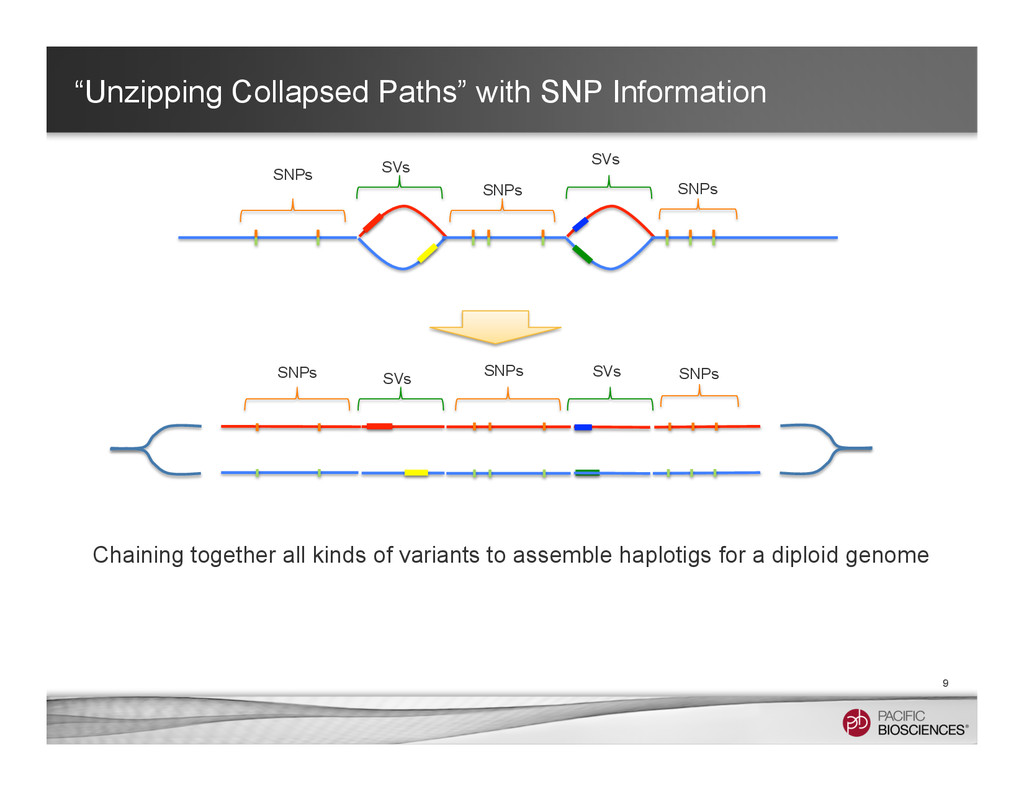
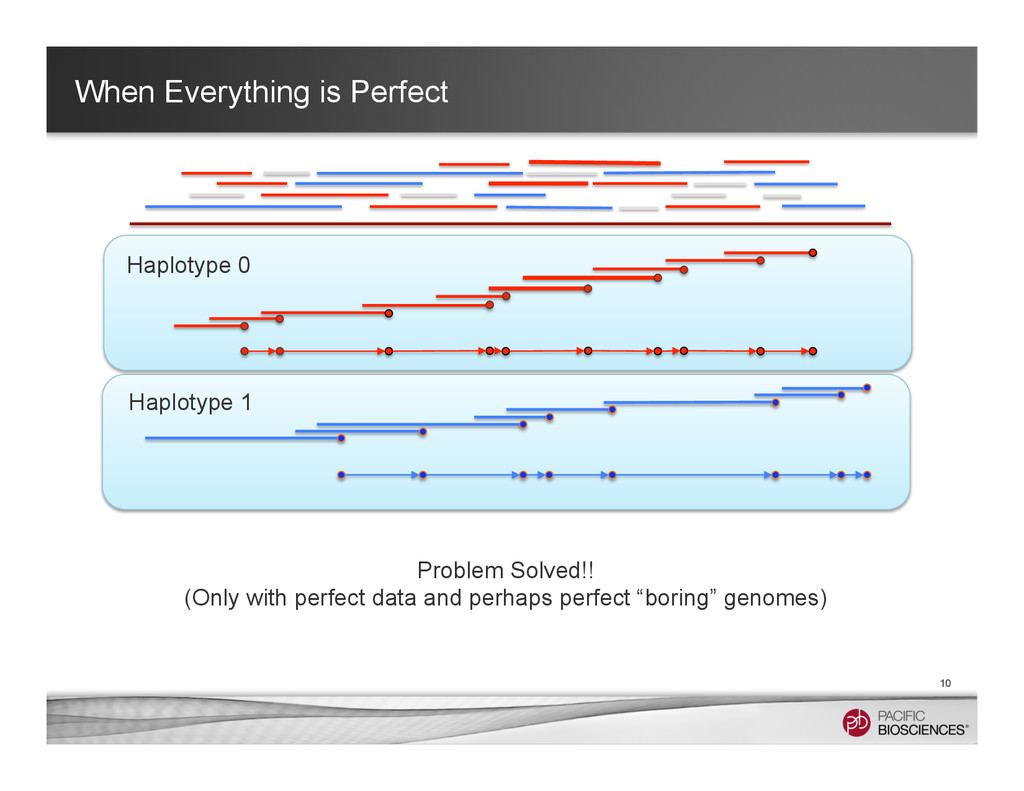
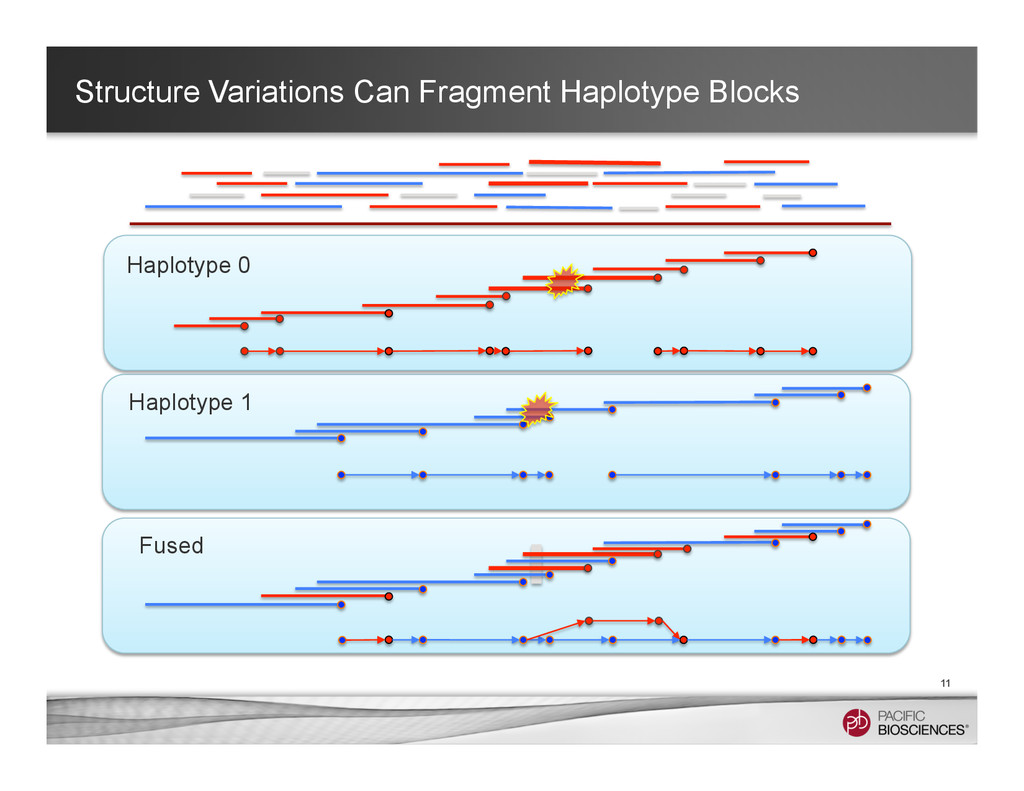
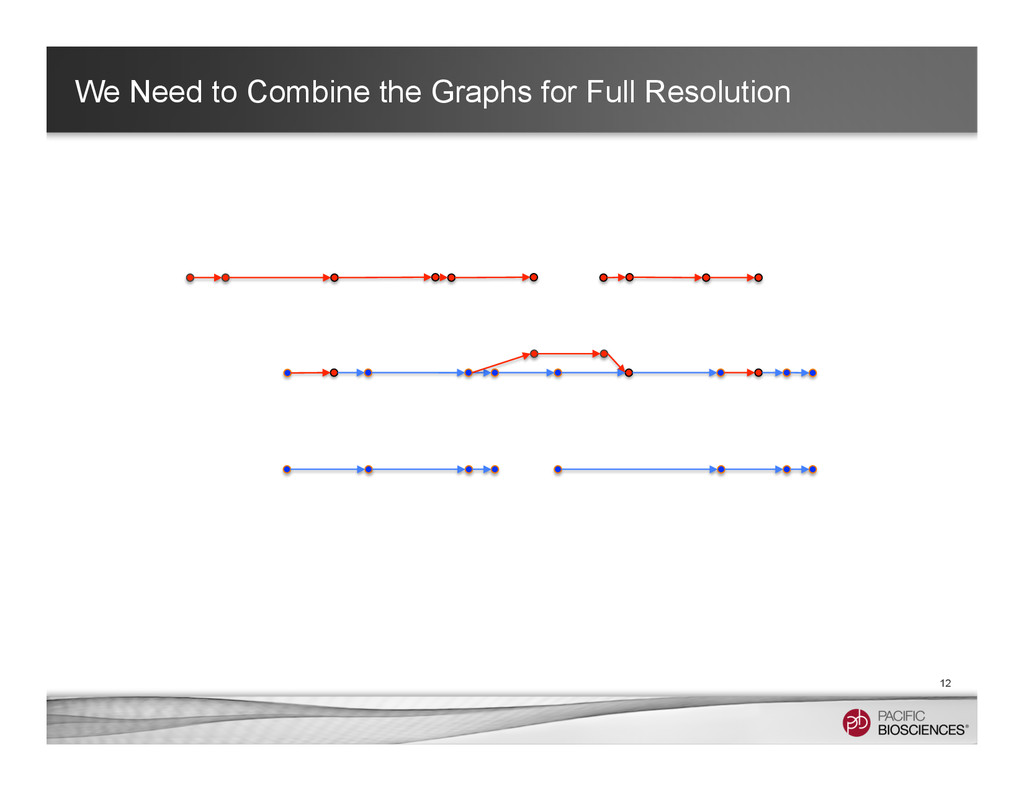
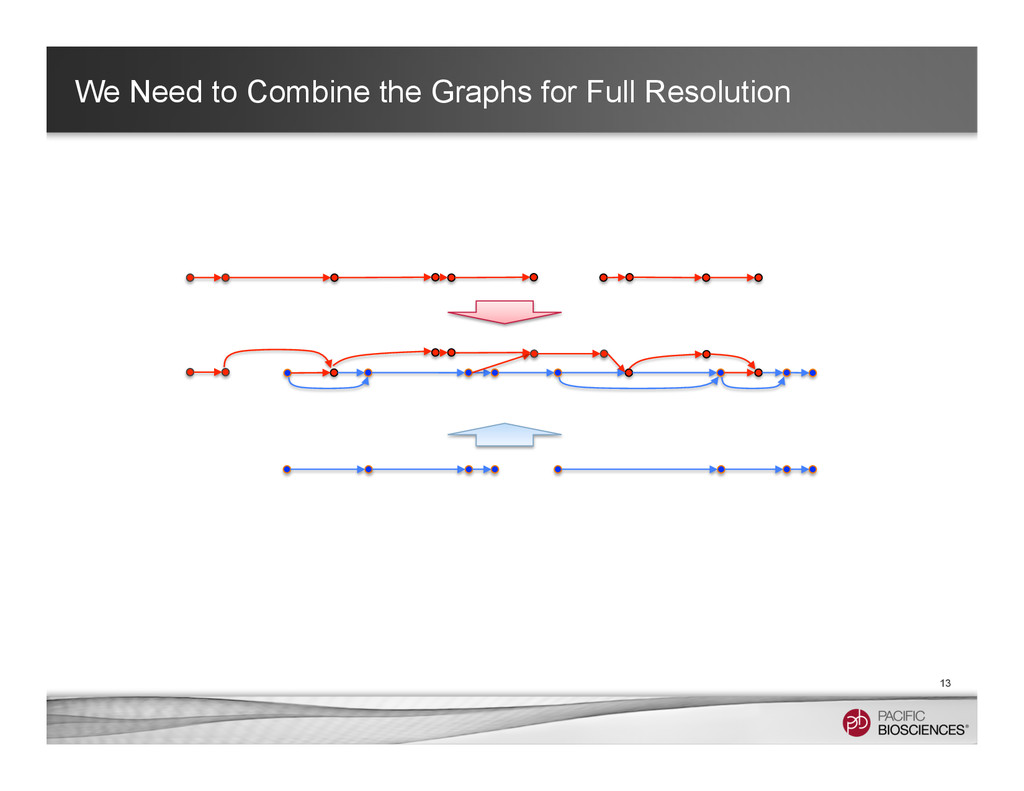
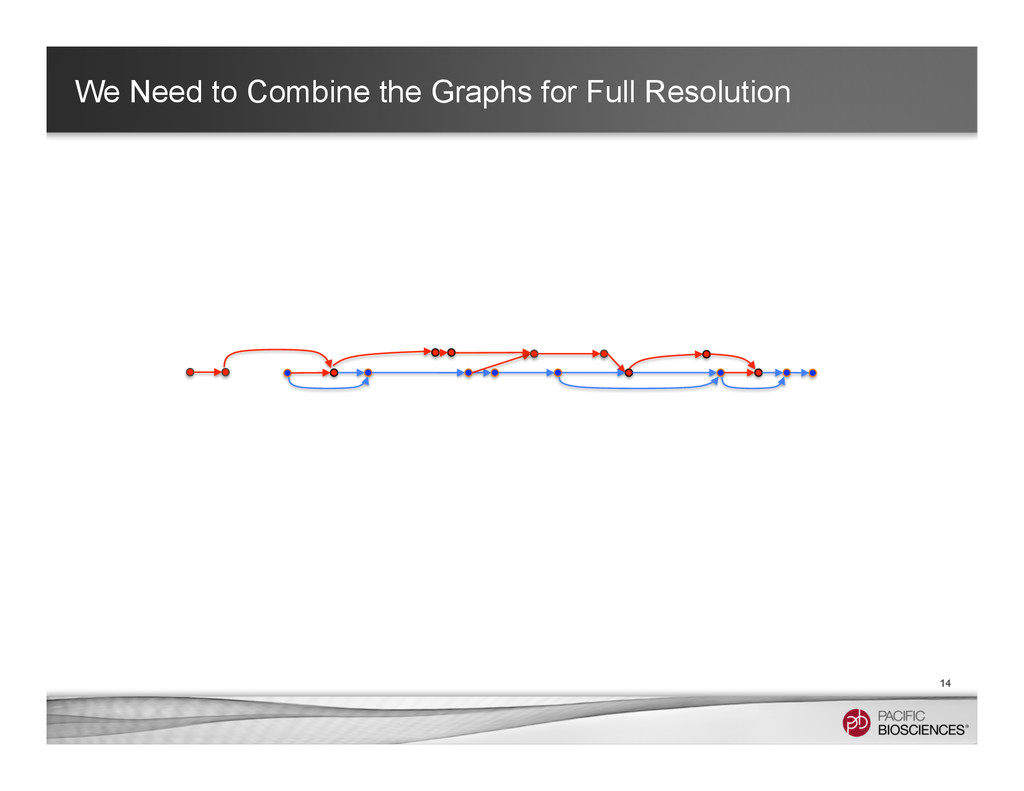
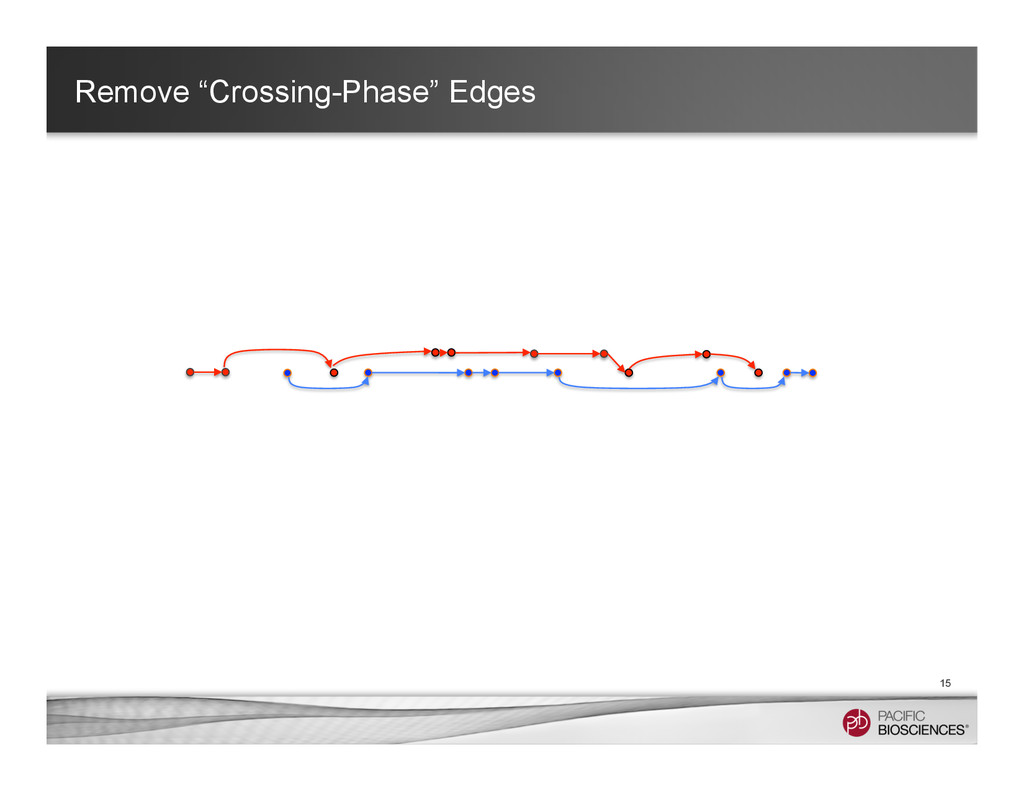
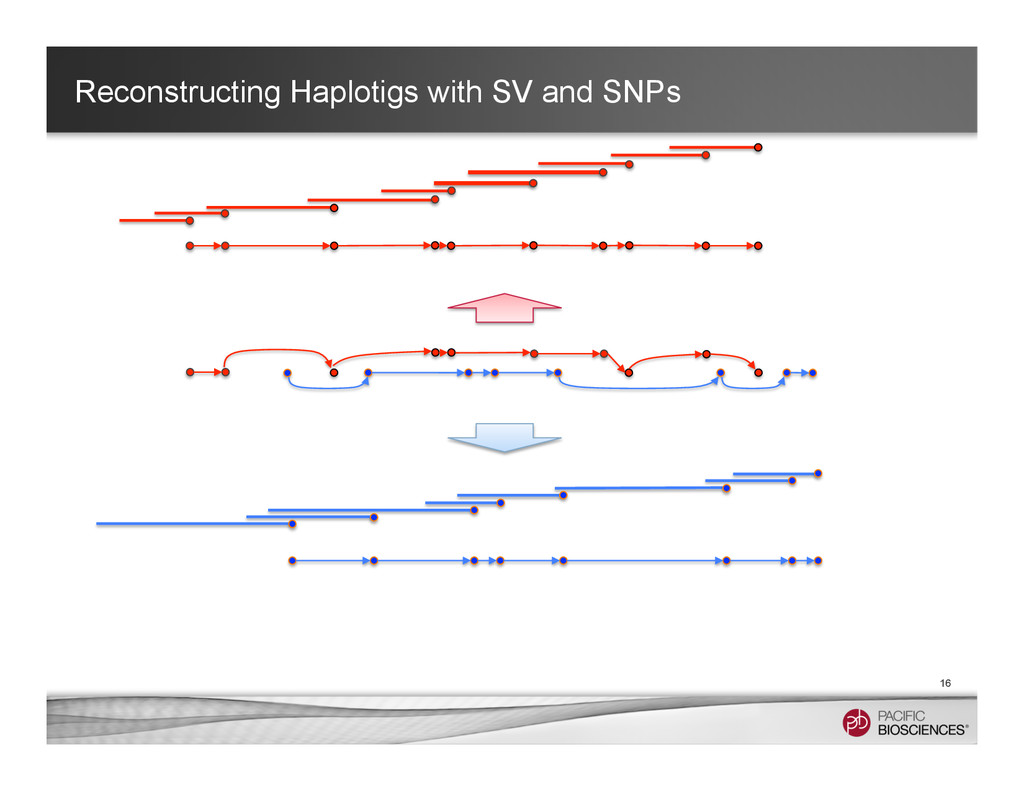
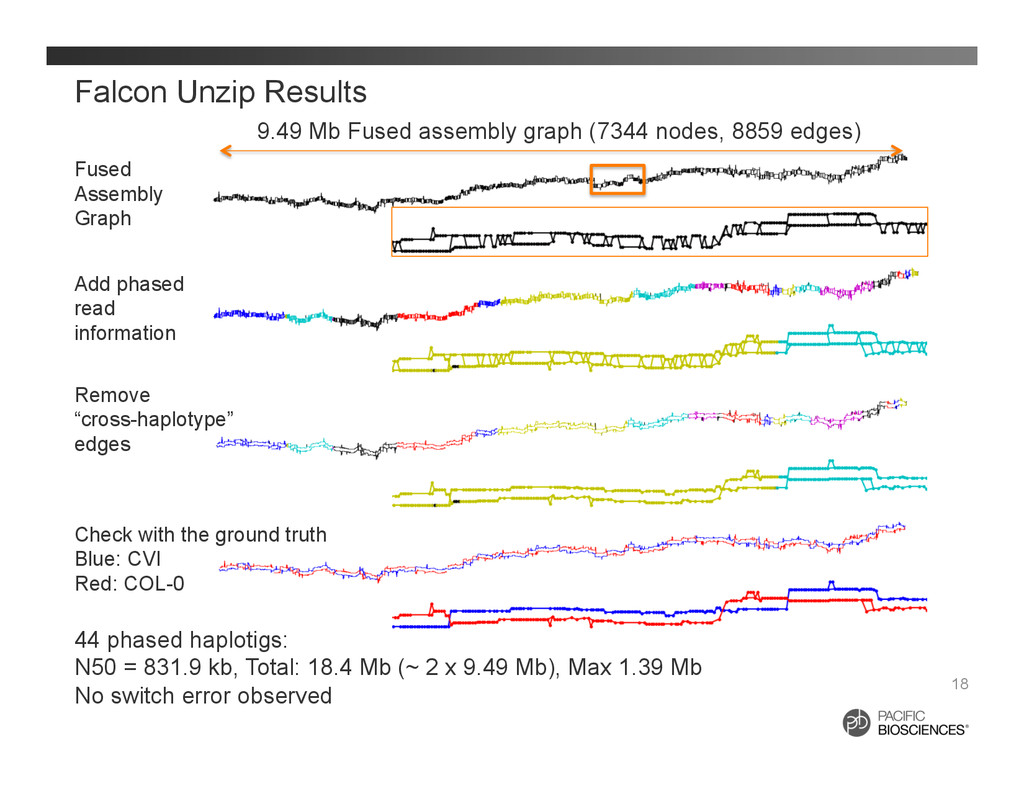
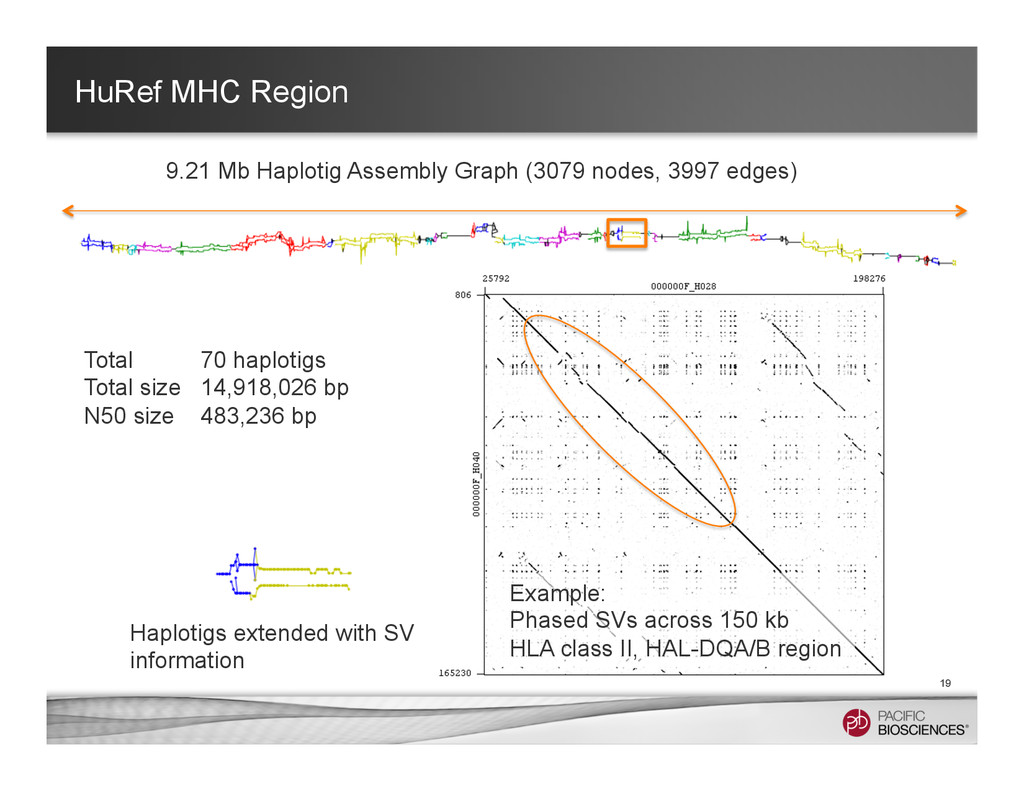
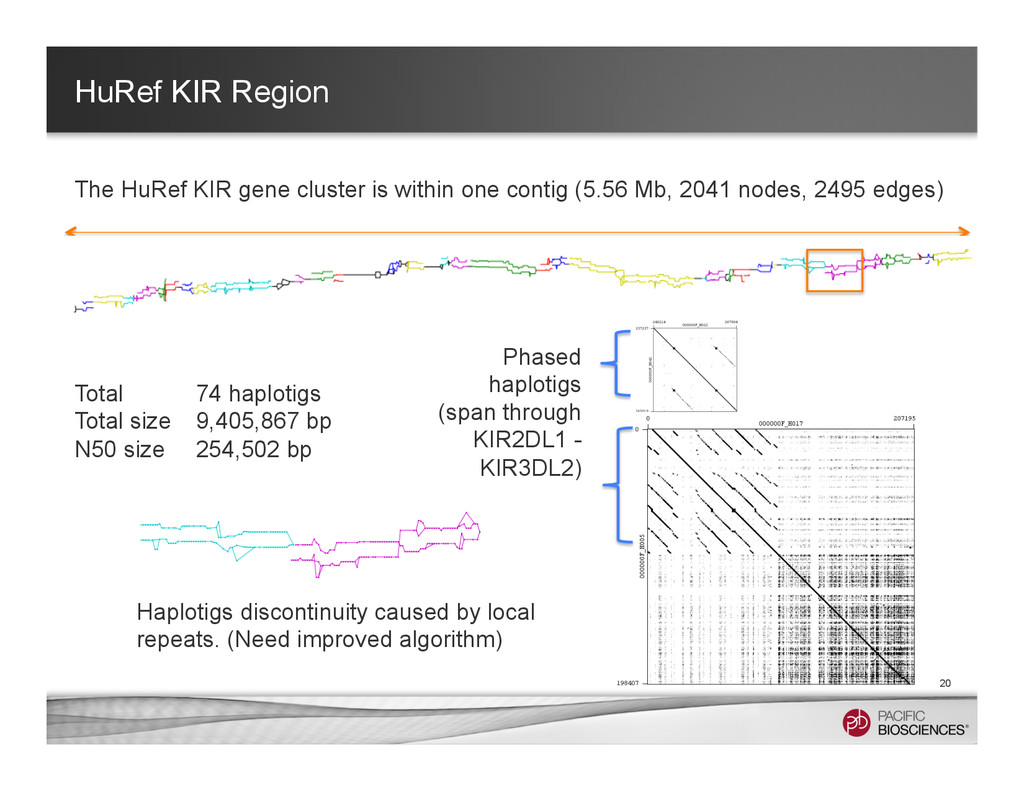
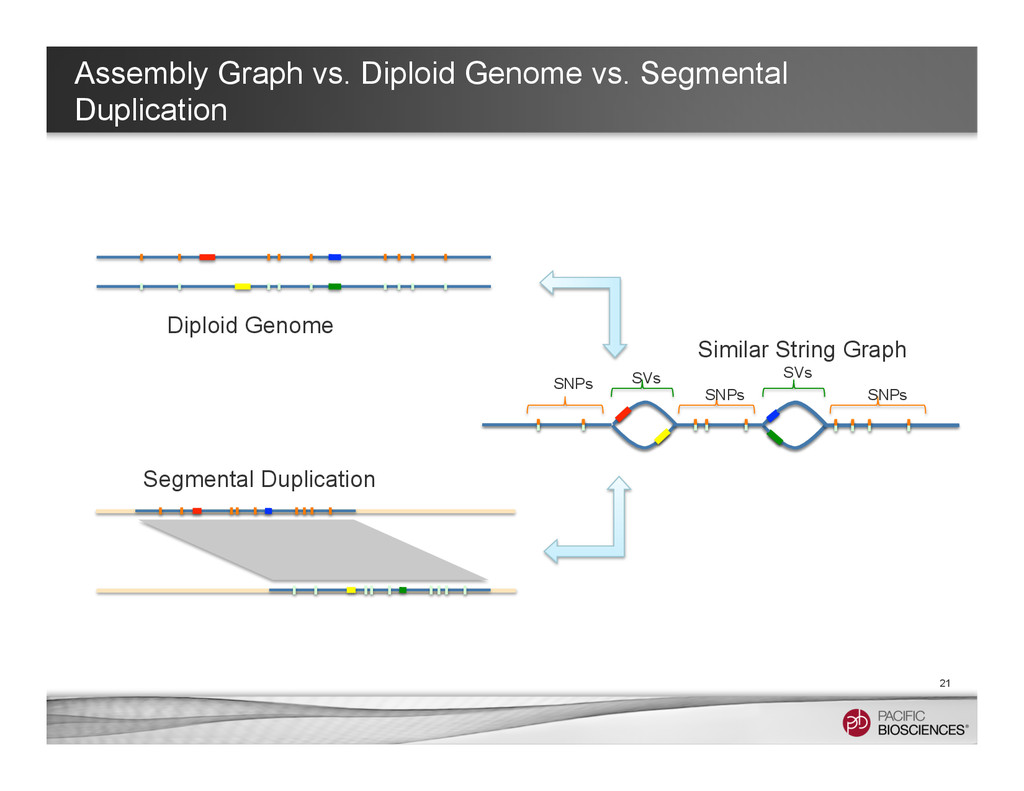
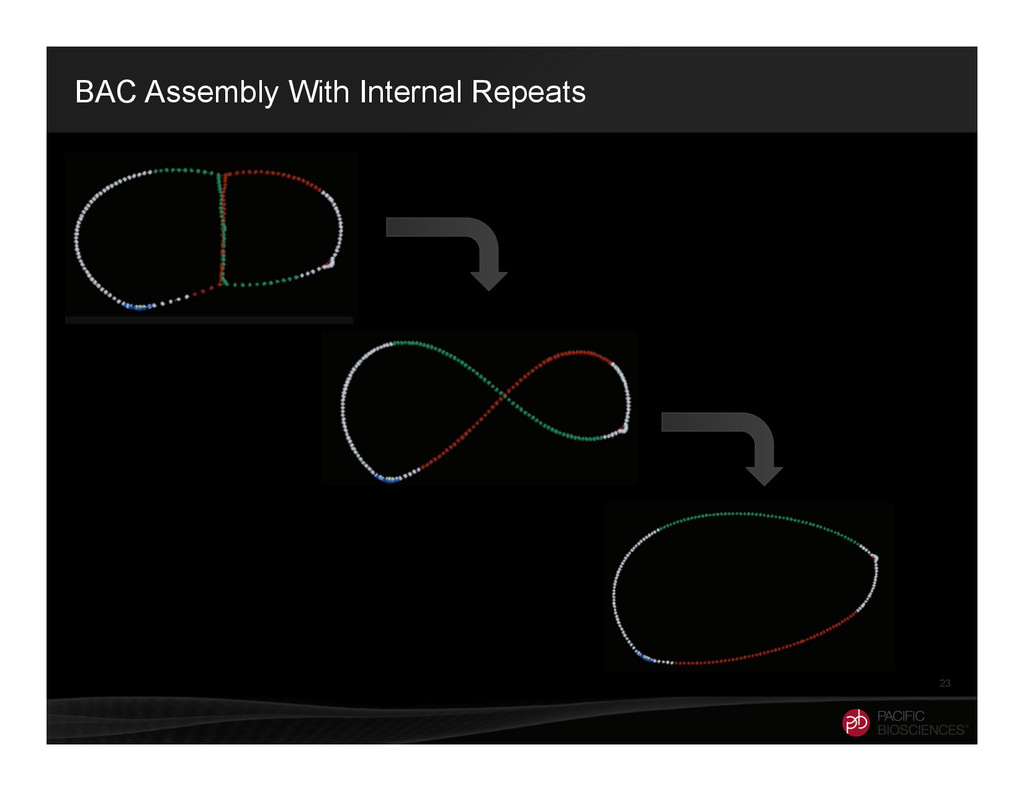
It is about how to do diploid genome assembly by using string graph formulation to construct haploptype graph and halplotigs.
The main point is it is possible to go beyond phasing SNPs only and only identify SNP level variations.
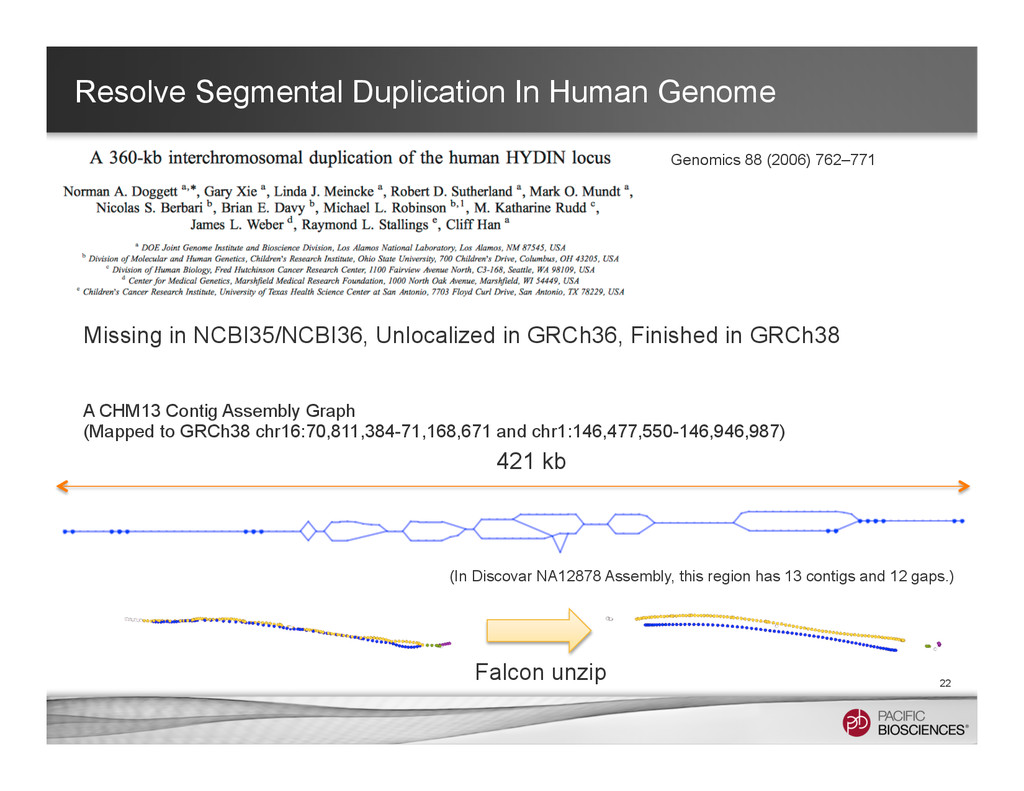
It is useful to reconstructing haplotype sequences including all variants with single molecule sequencing with PacBio platform to understand the biology of genome. We hope this sheds light on revealing interesting biology by sequence real diploid genomes routinely without constructing inbred lines for sequencing.
I also make a couple videos for fun to demonstrate the process:
diploid assembly to haplotig example (long version)
https://www.youtube.com/watch?v=vwSyD31eahI
diploid assembly to haplotig example (short version)
https://www.youtube.com/watch?v=yC1ujdLUT7Q
Genome Assembly Problem and 1-D Cosmology
https://www.youtube.com/watch?v=rv9VV-sG4rs
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}