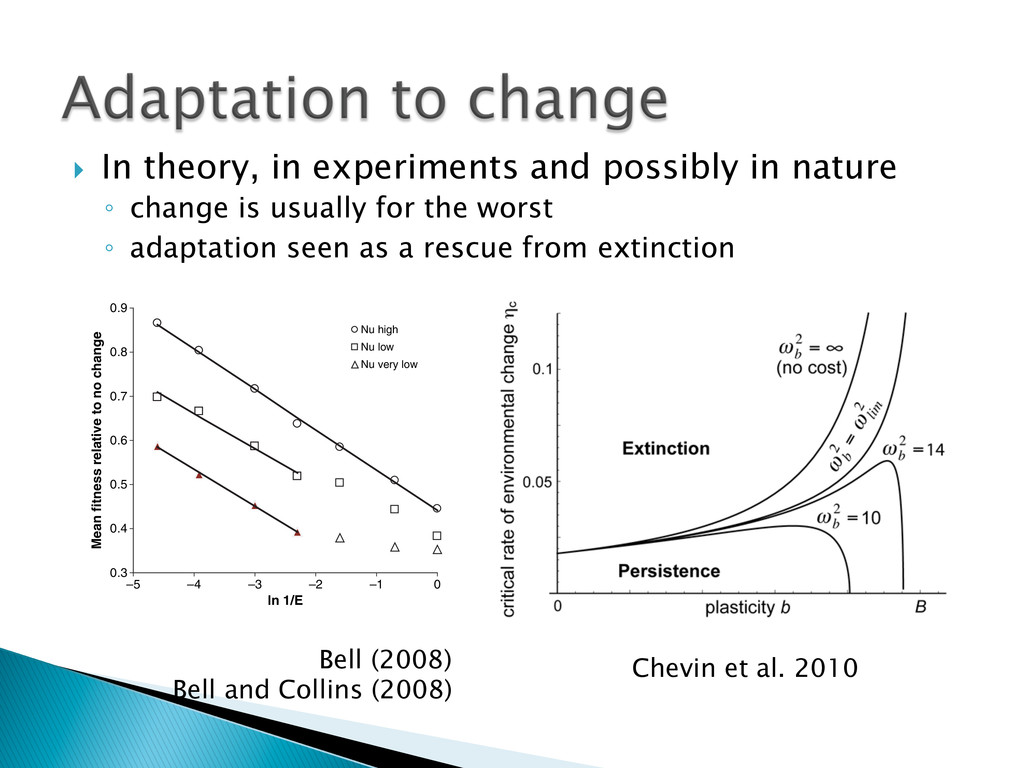
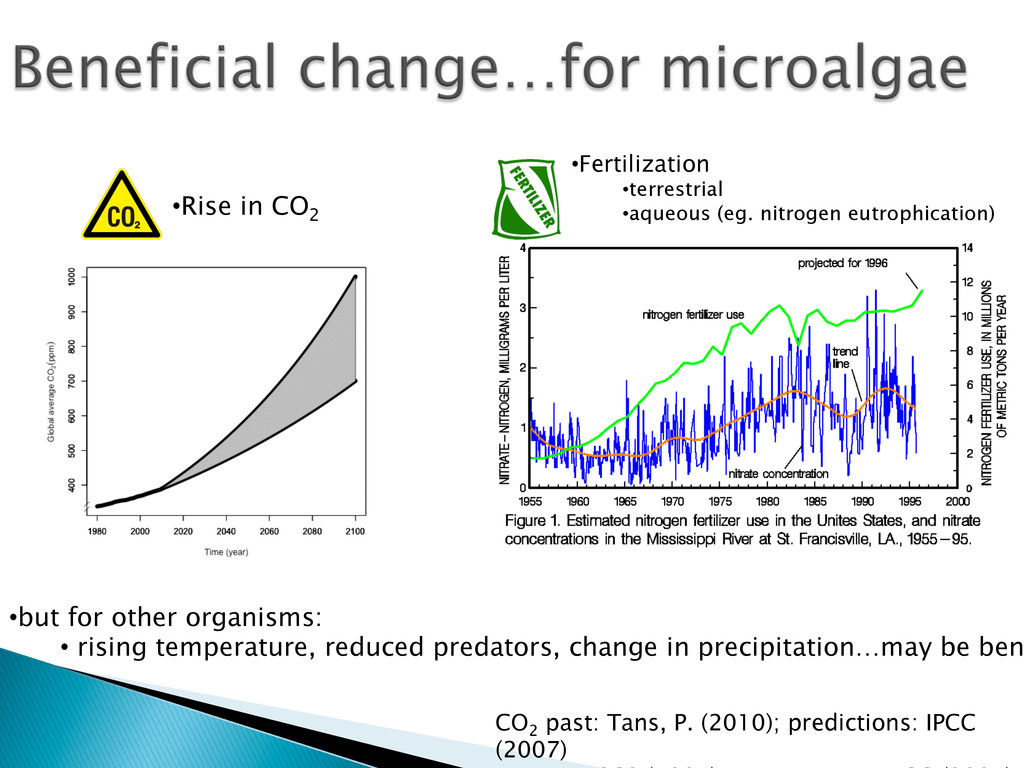
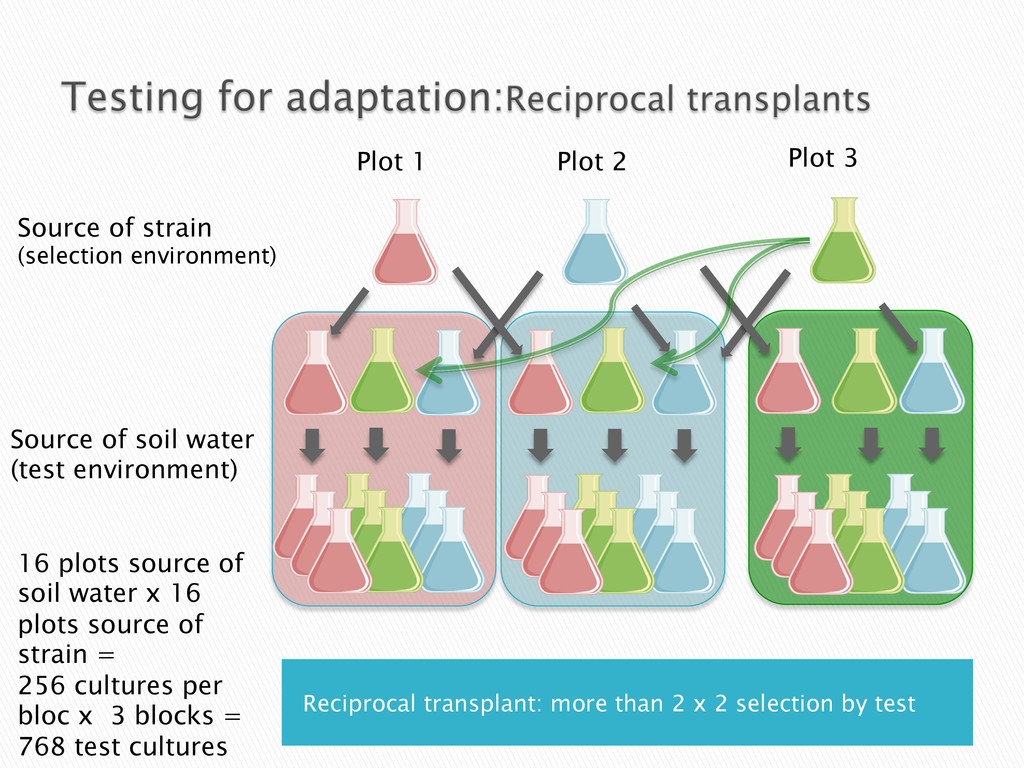
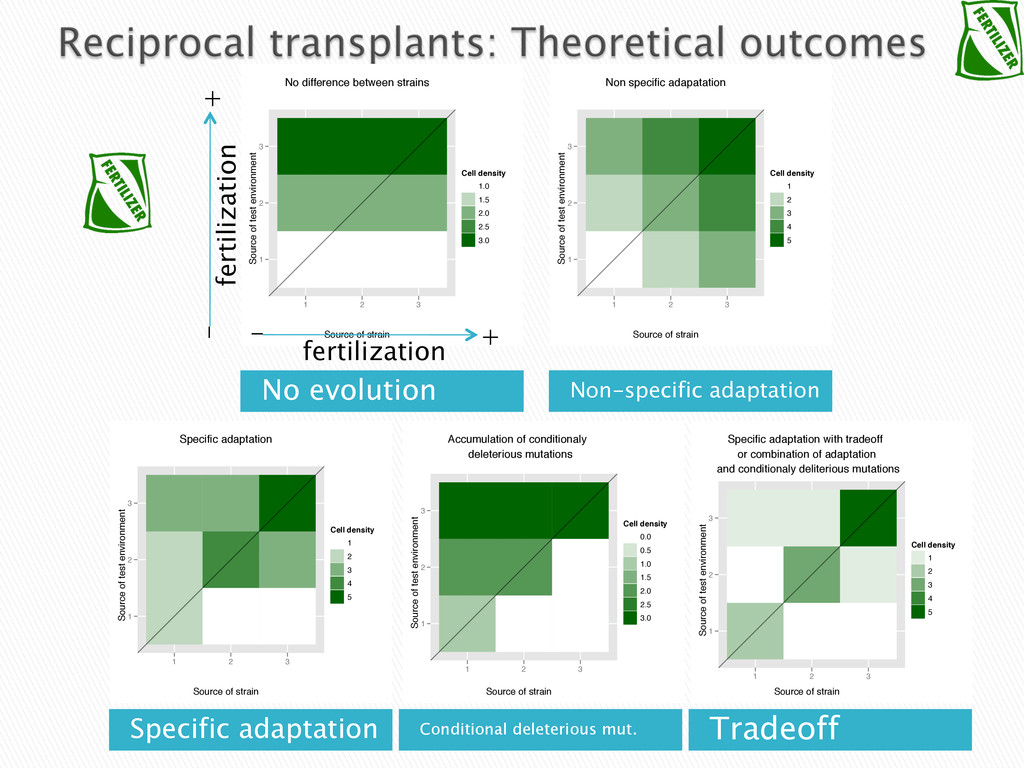
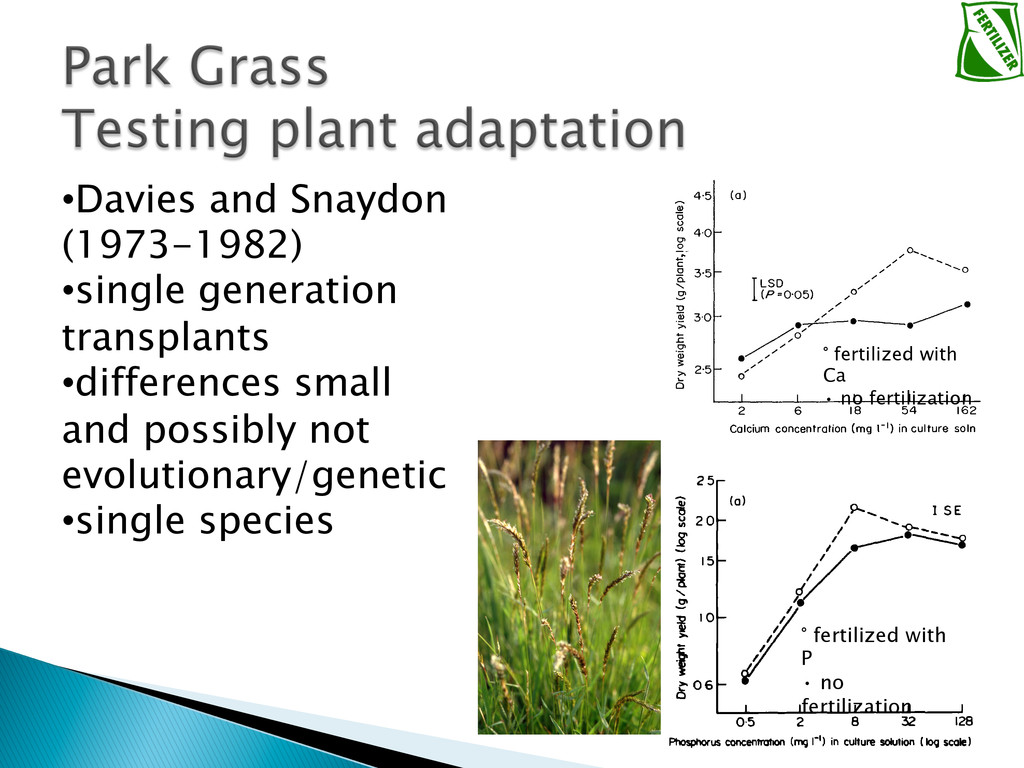
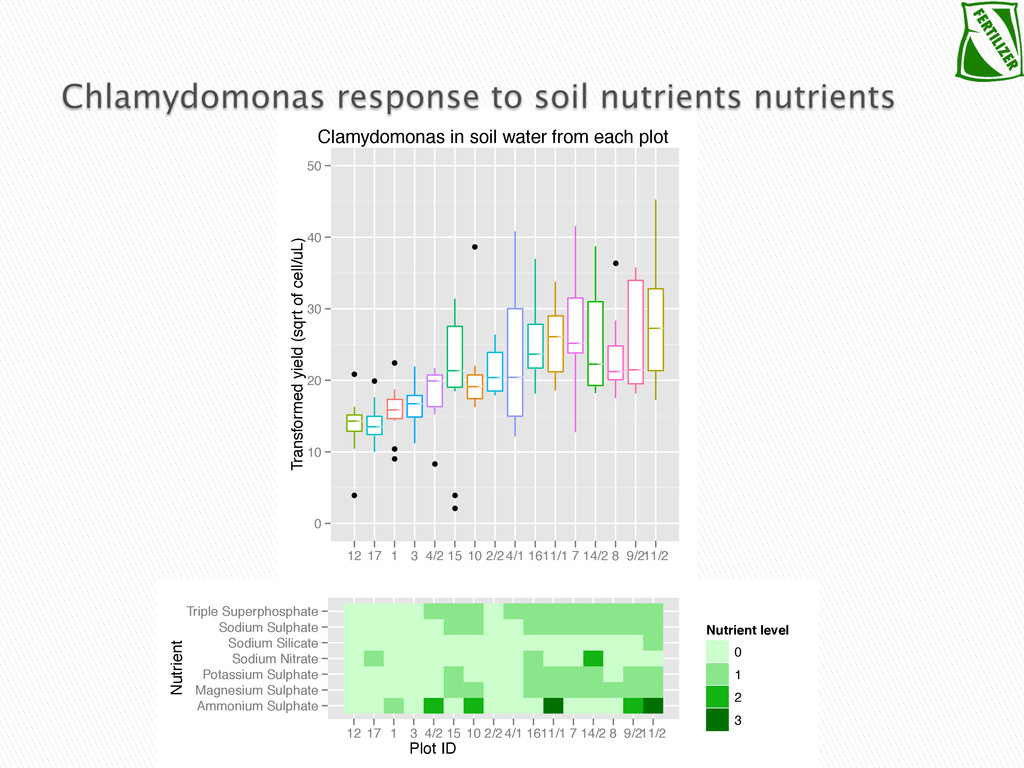
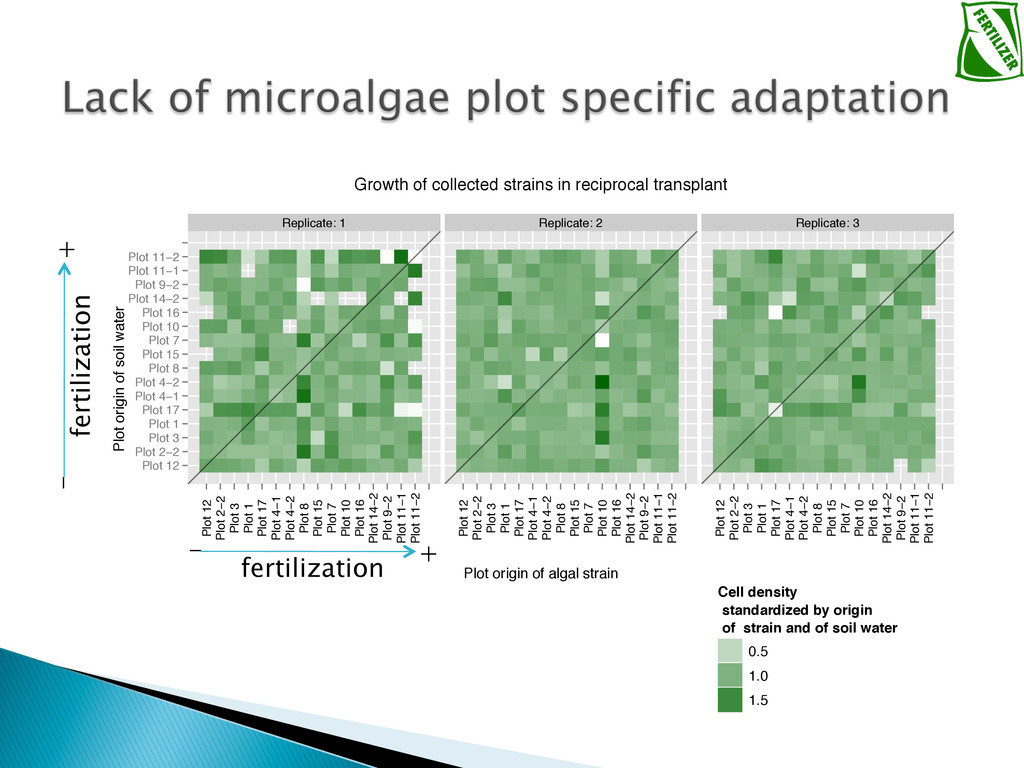
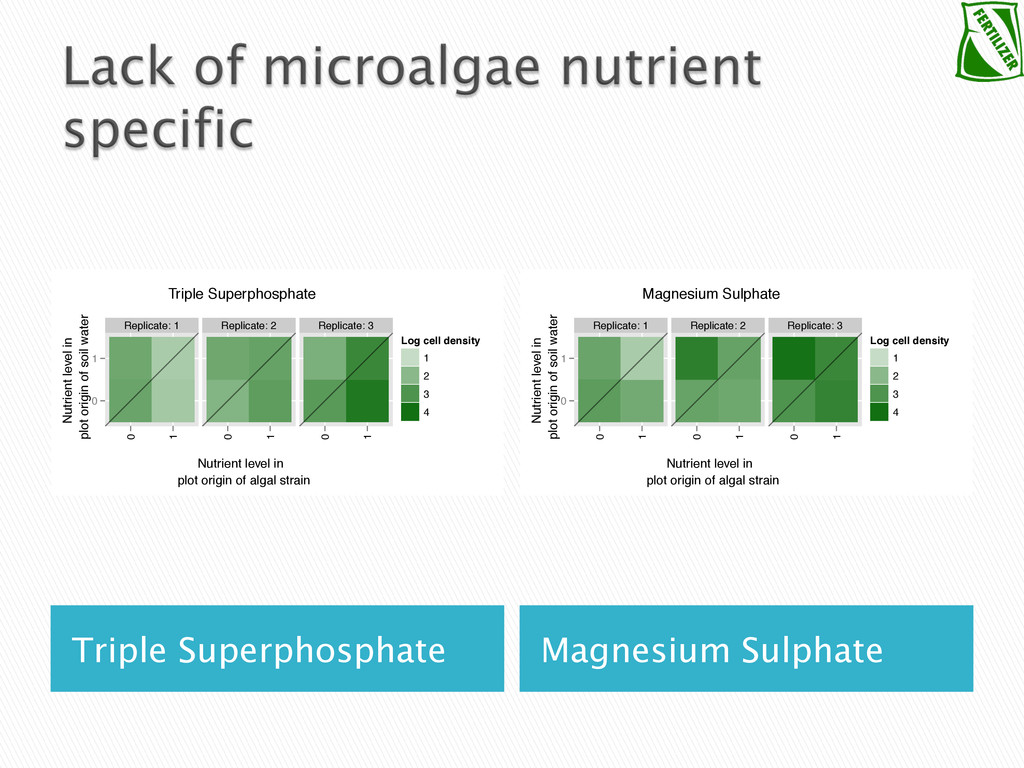
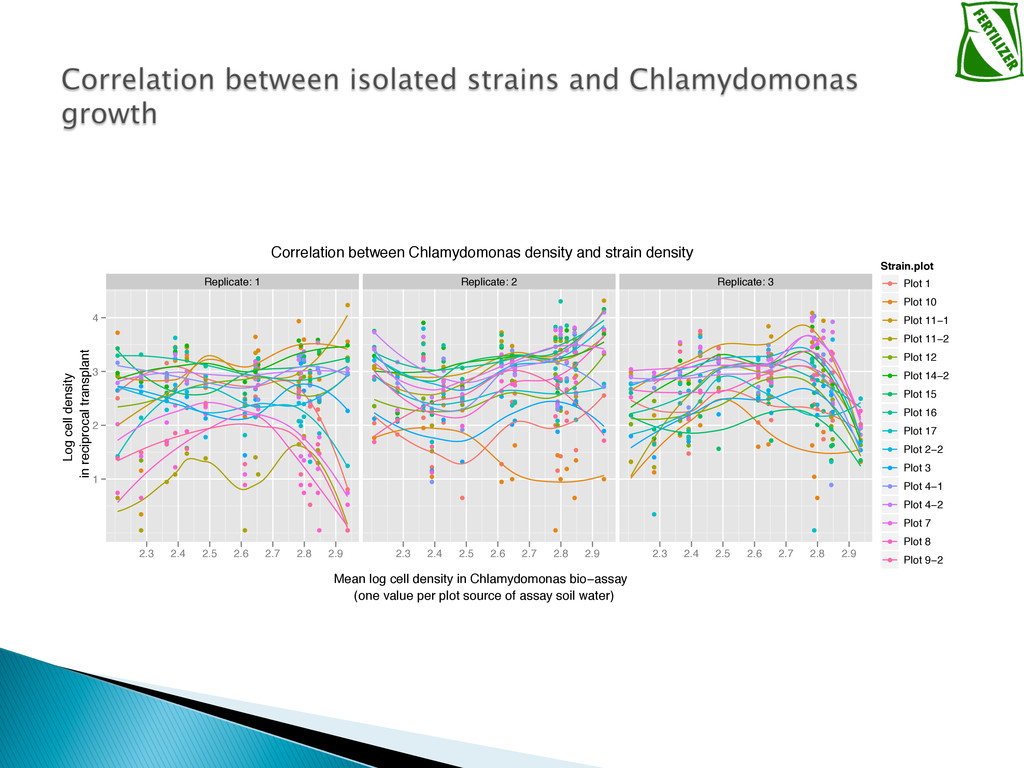
Experimental evolution has predominantly focused on survival and adaptation of organisms subject to stressful changes in their environment. However, in many cases, environmental change shifts conditions towards the optimum of a species. Current theory may not be well suited to predicting the response of organisms to this type of changes. We investigate the adaptation of soil micro-algae to the addition of a variety of fertilizers. We isolated and tested soil micro-algae from plots of the Park Grass Experiment at Rothamsted Research, which have been subject to fertilization treatment continuously for over 150 years. This research will test evolutionary theory in new contexts and will help predict the response of organisms benefiting from global changes, including rising CO2, eutrophication, increased nitrogen availability and rising temperatures.
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}