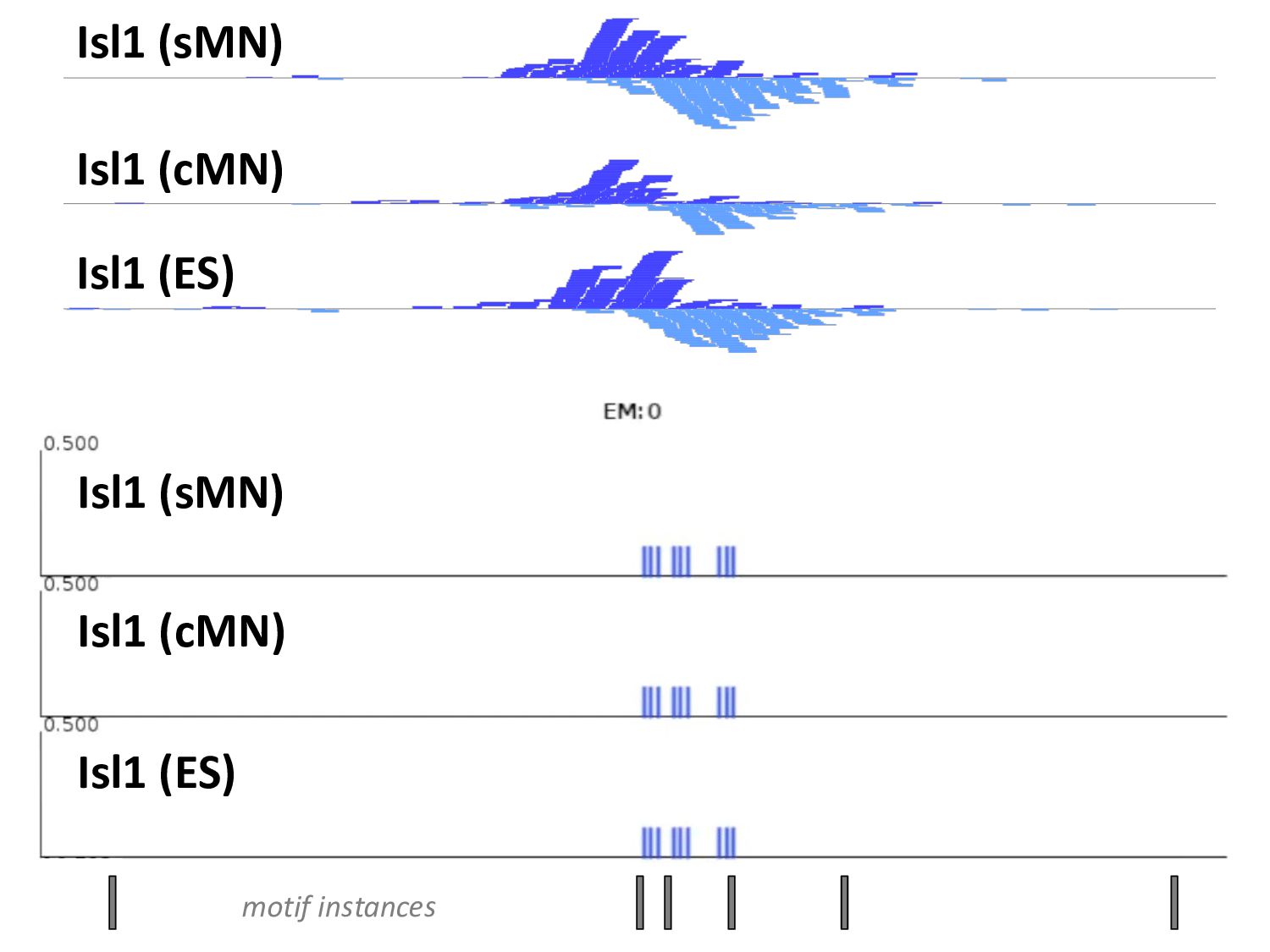
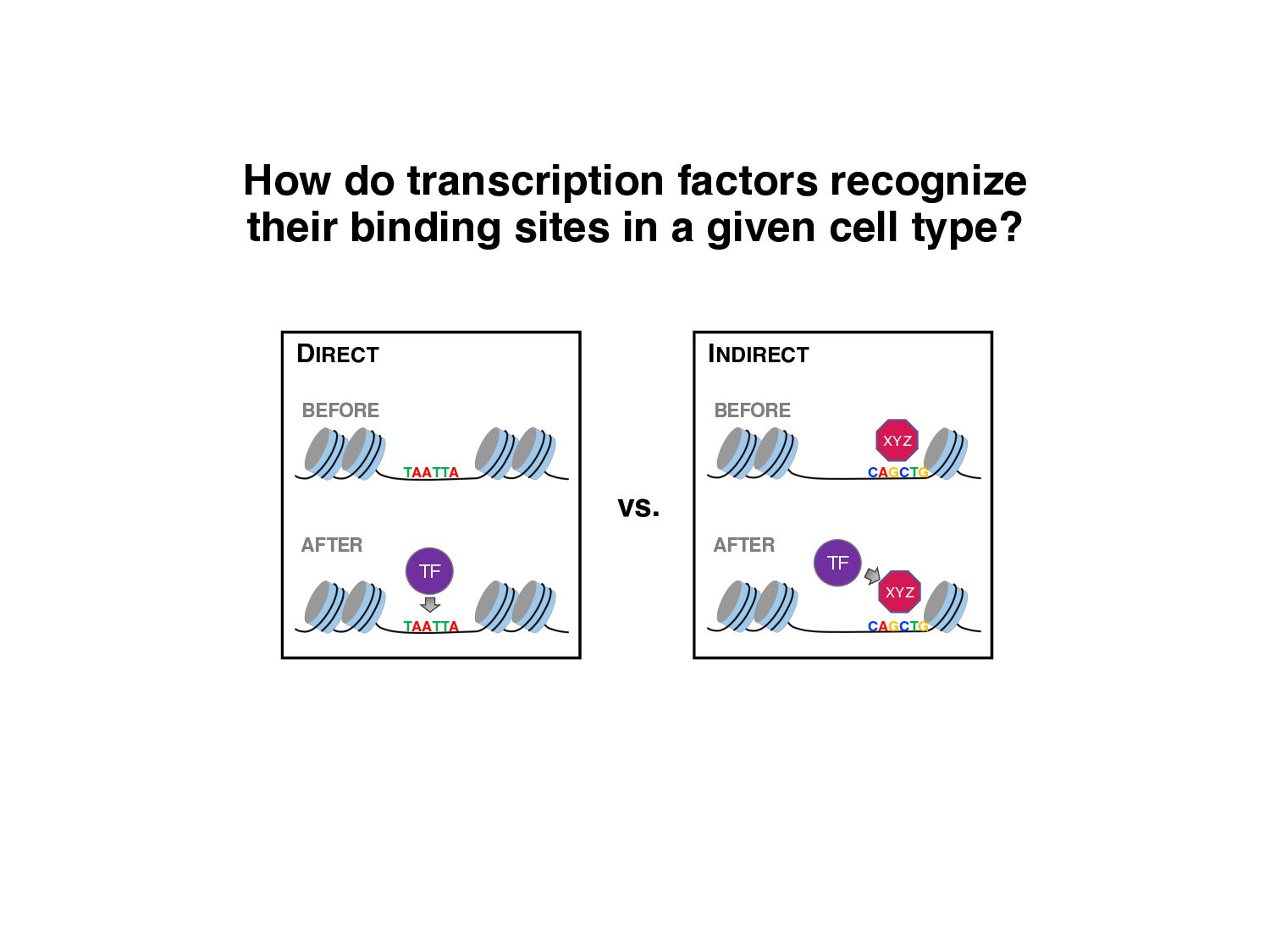
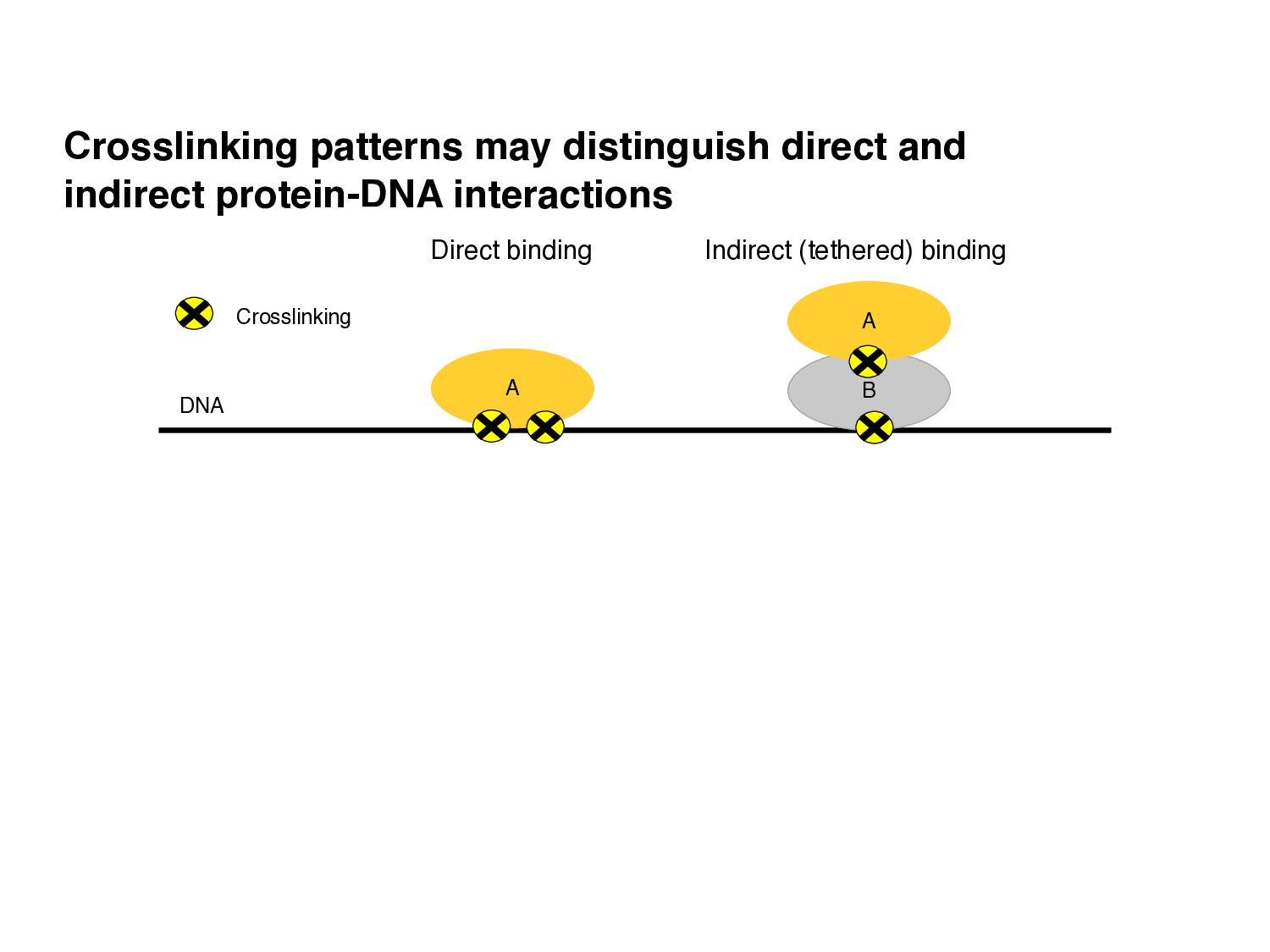
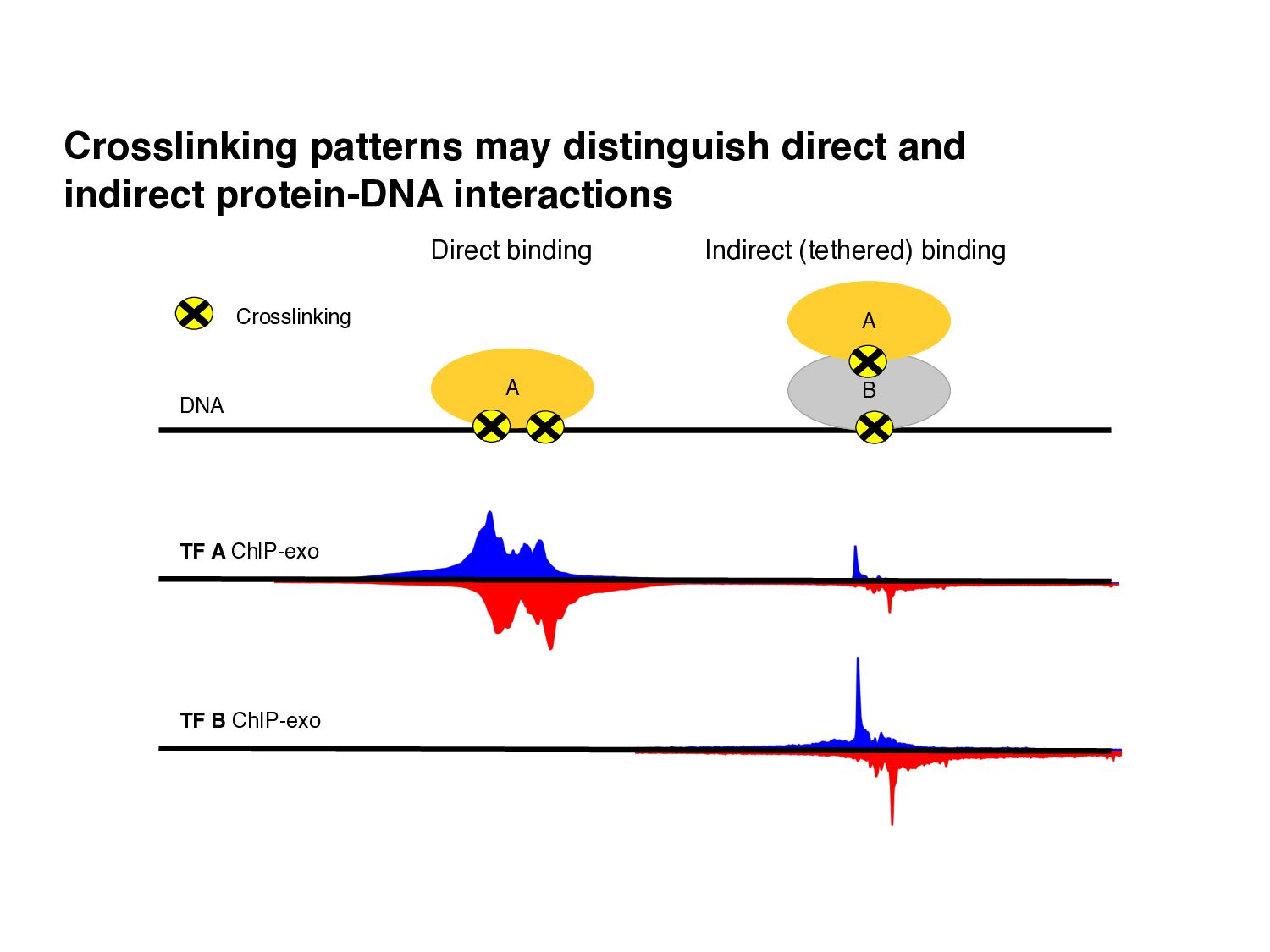
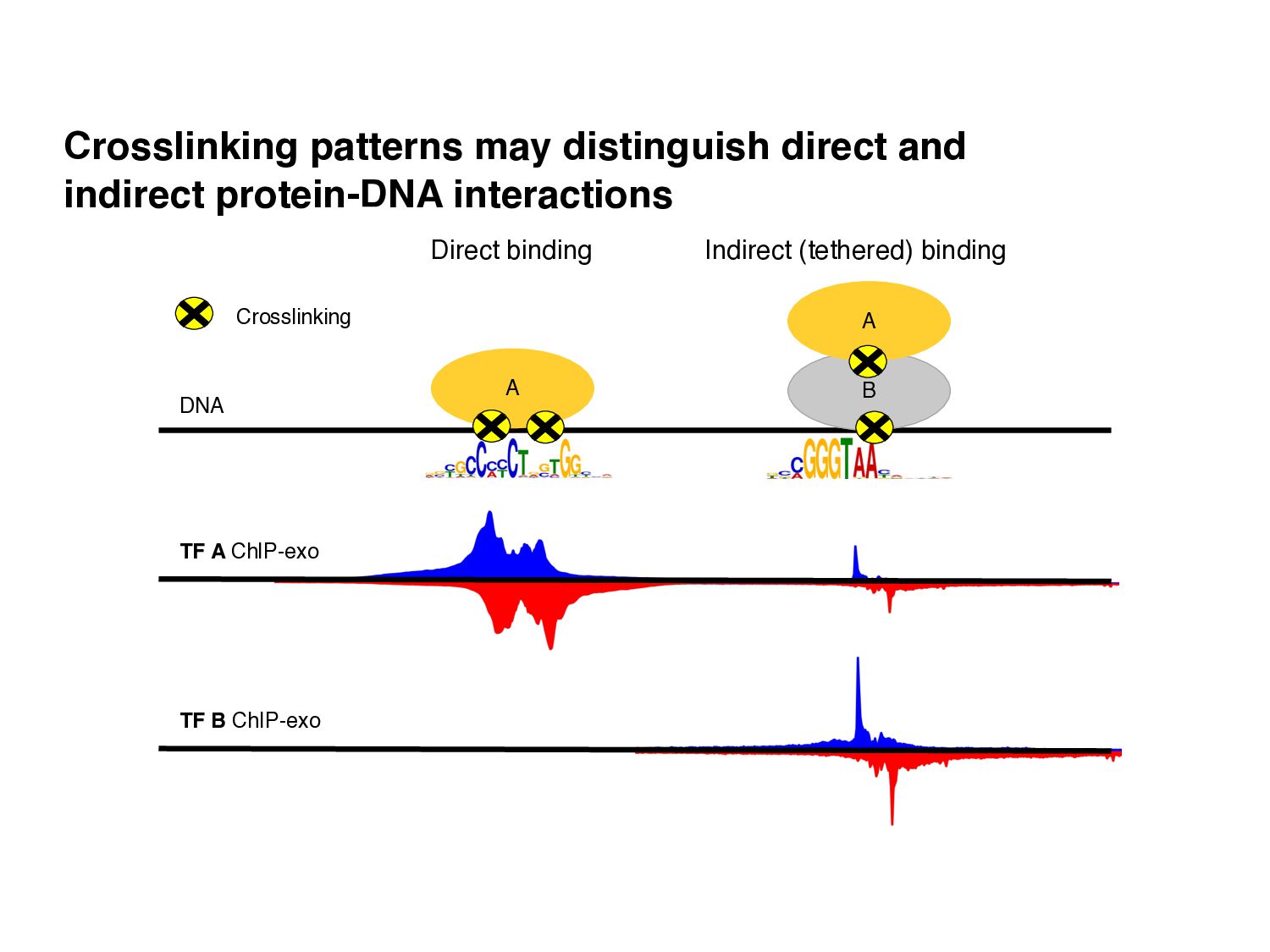
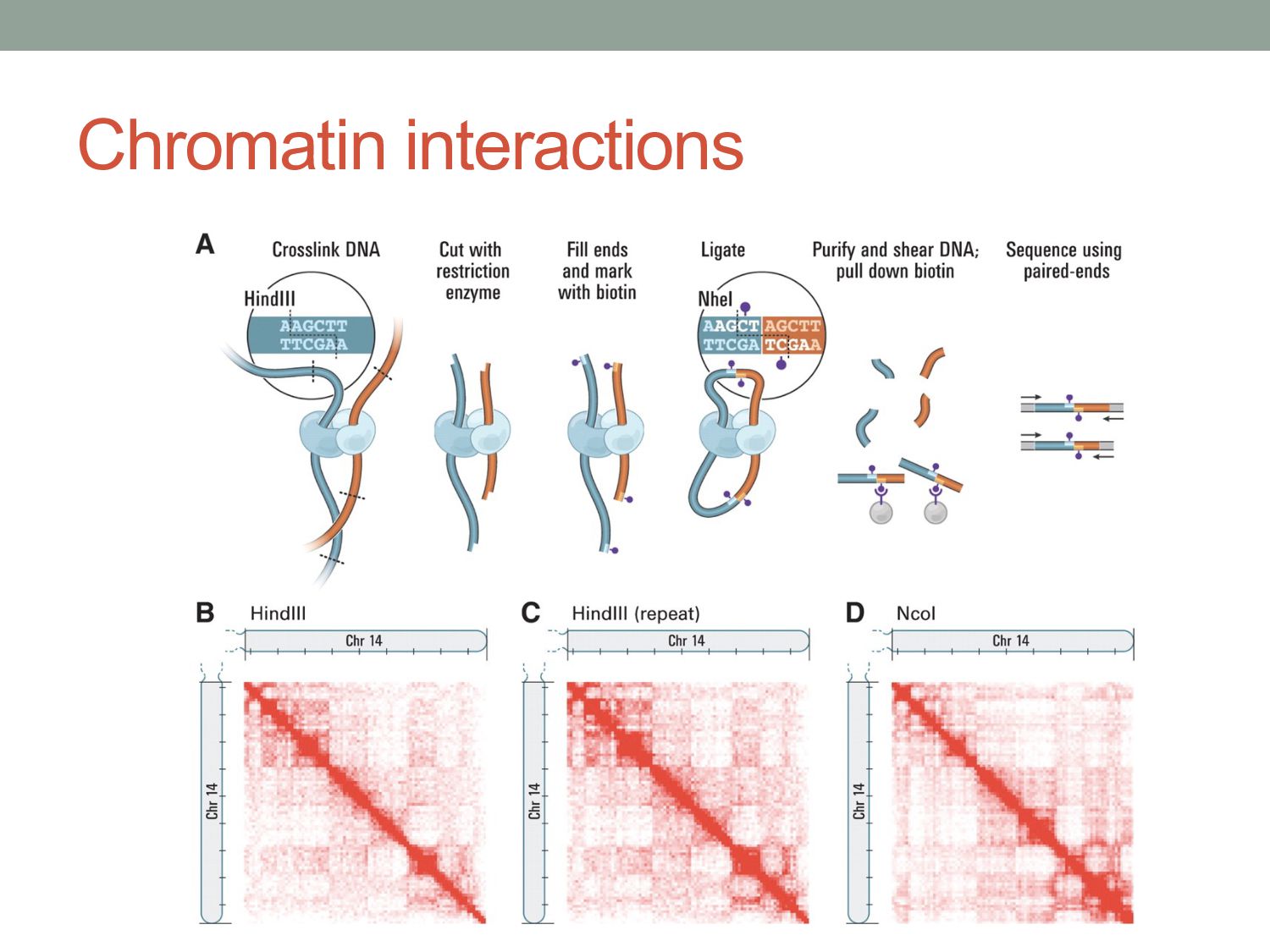
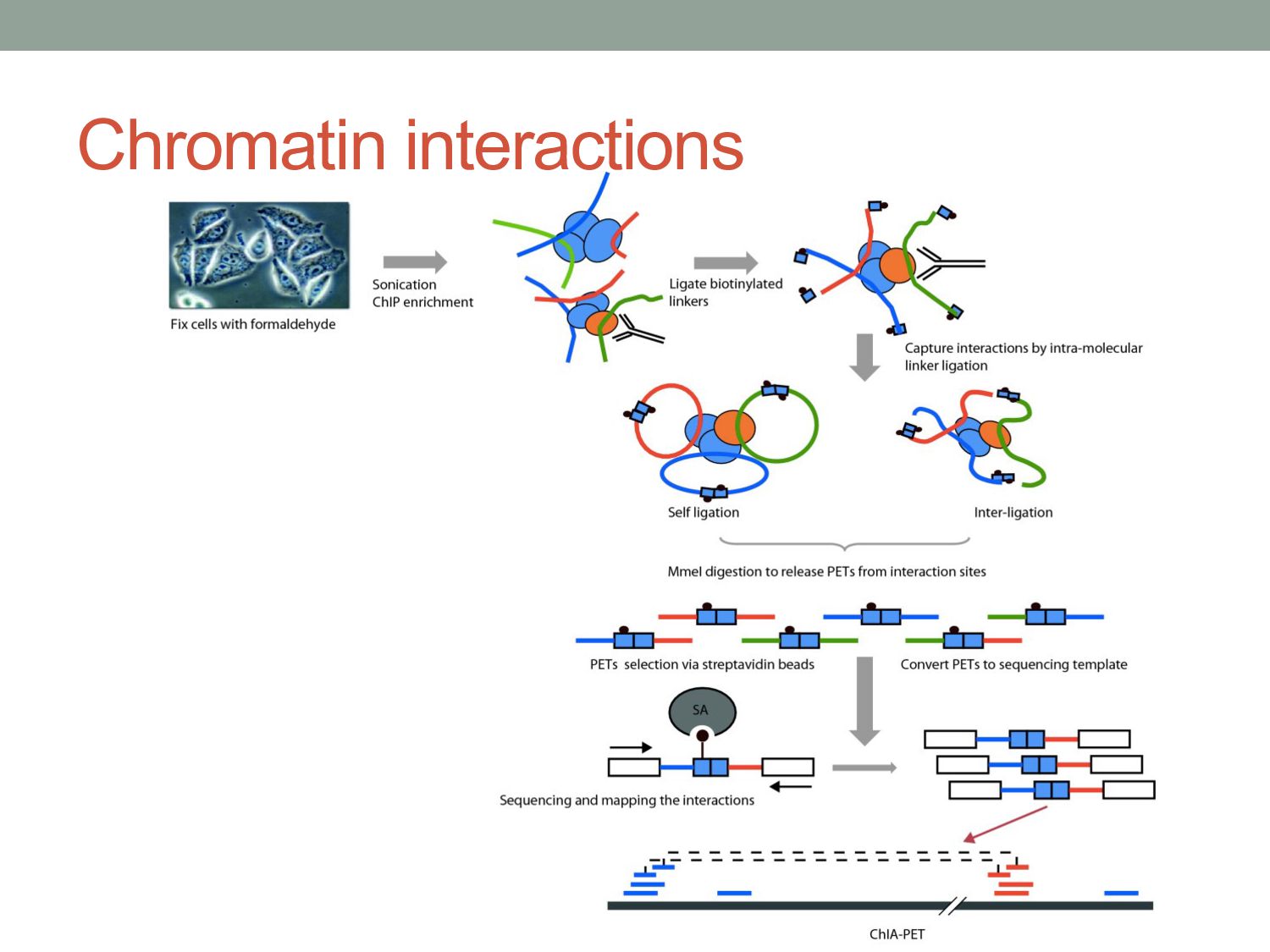
of protein-DNA interaction from ChIP-seq data? • How can we find differences in regulatory signals across conditions? • Can we determine whether proteins are bound directly or indirectly to DNA? • Can we assay three-dimensional interactions in chromatin?
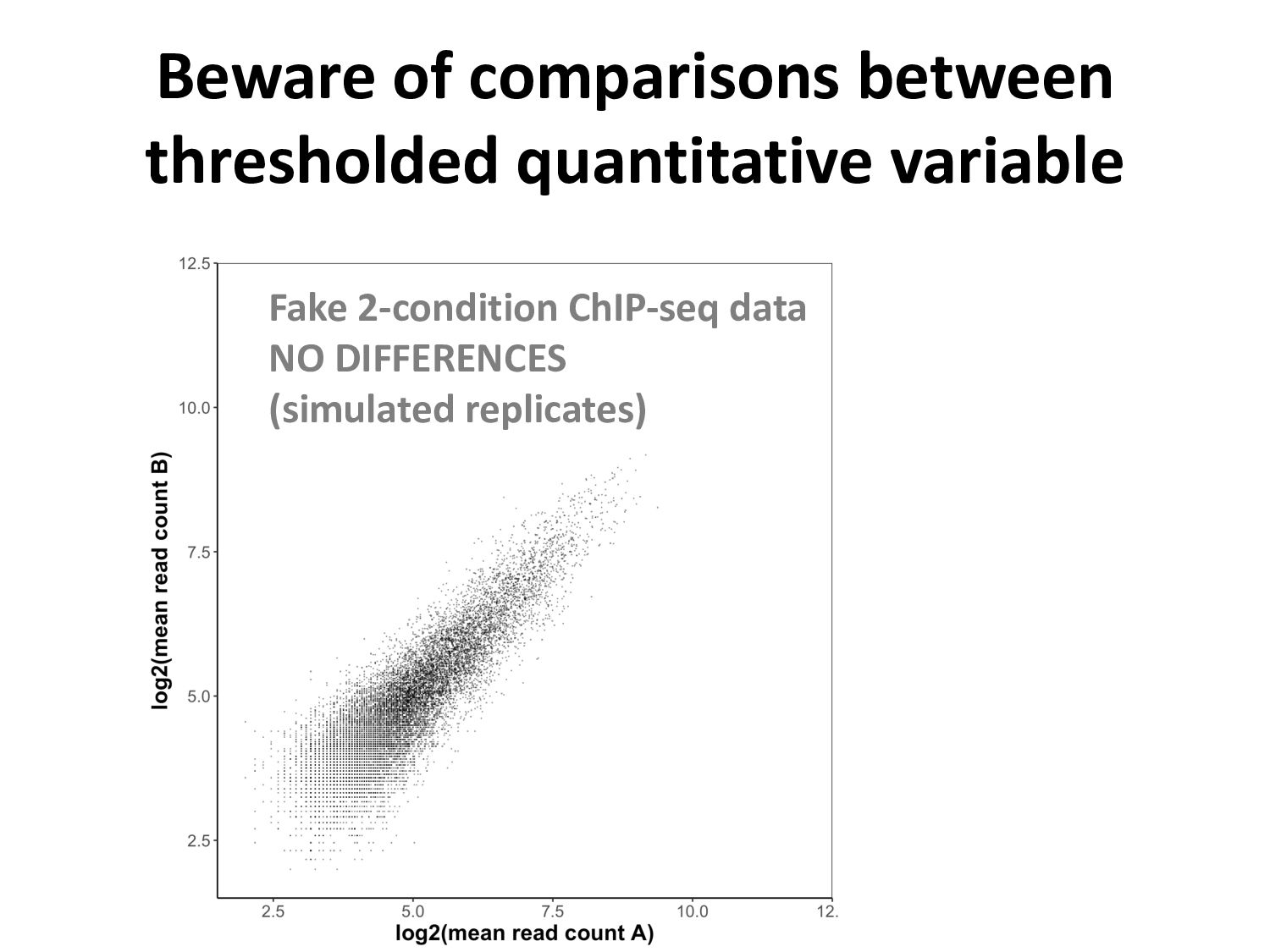
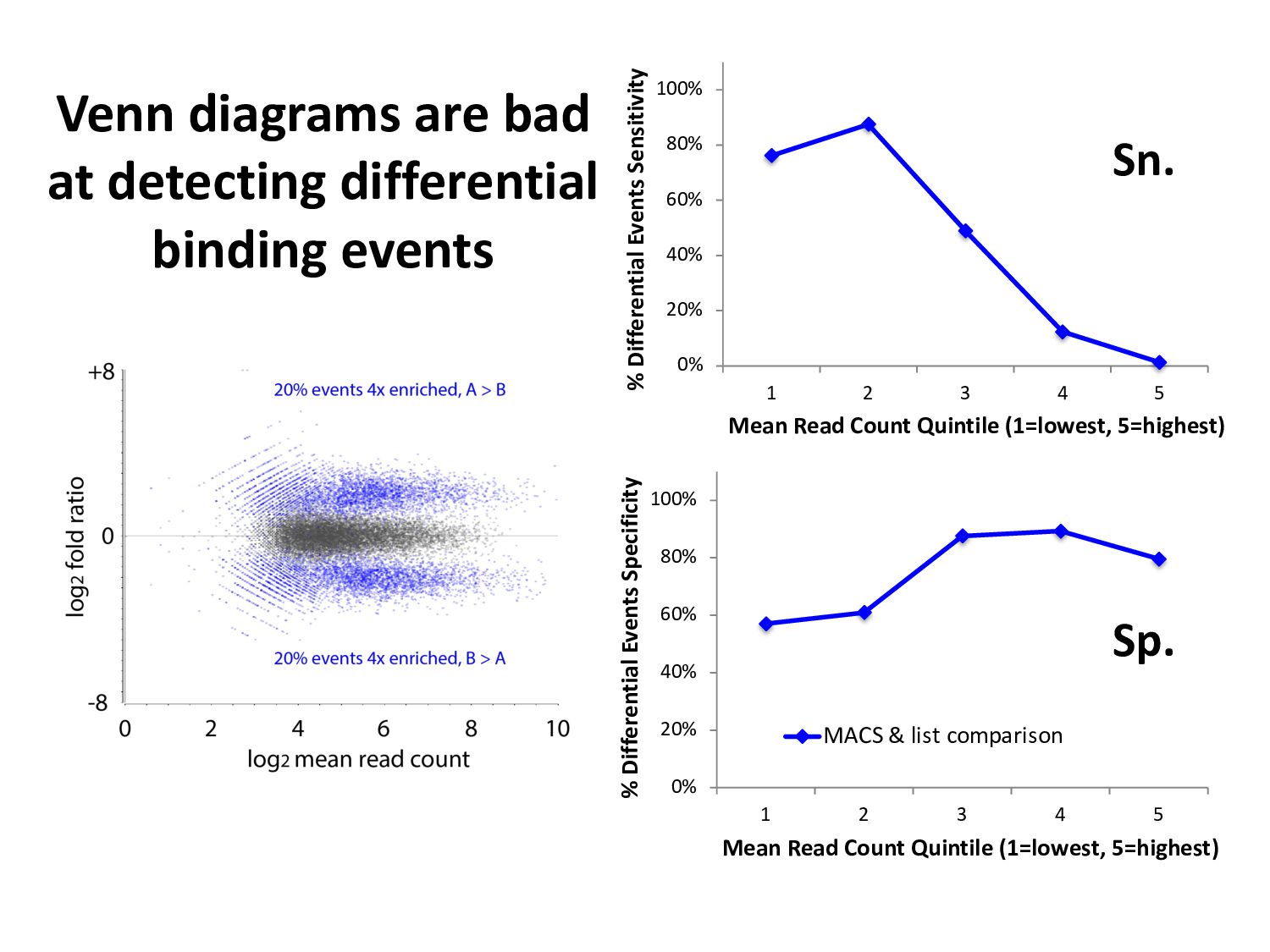
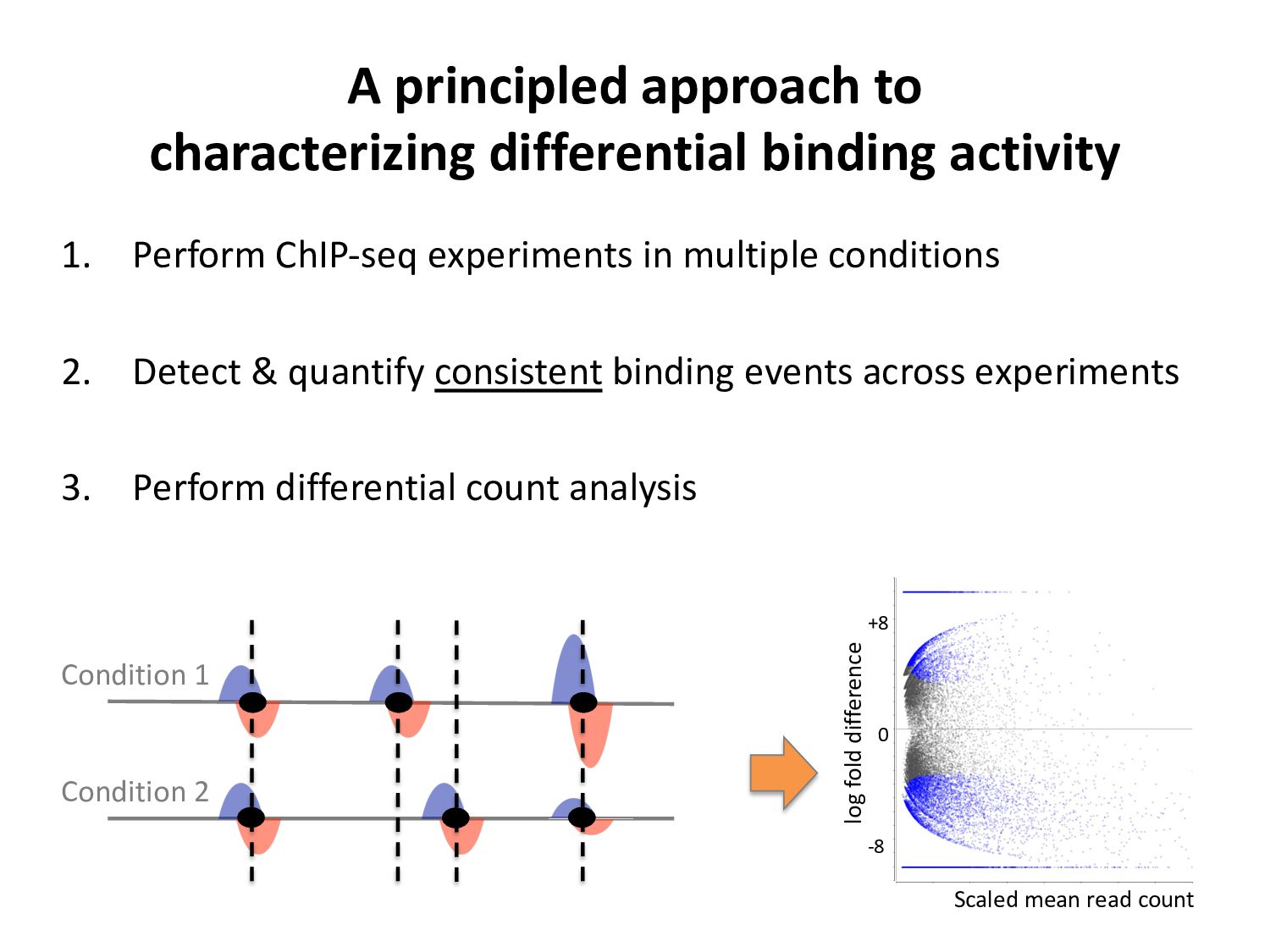
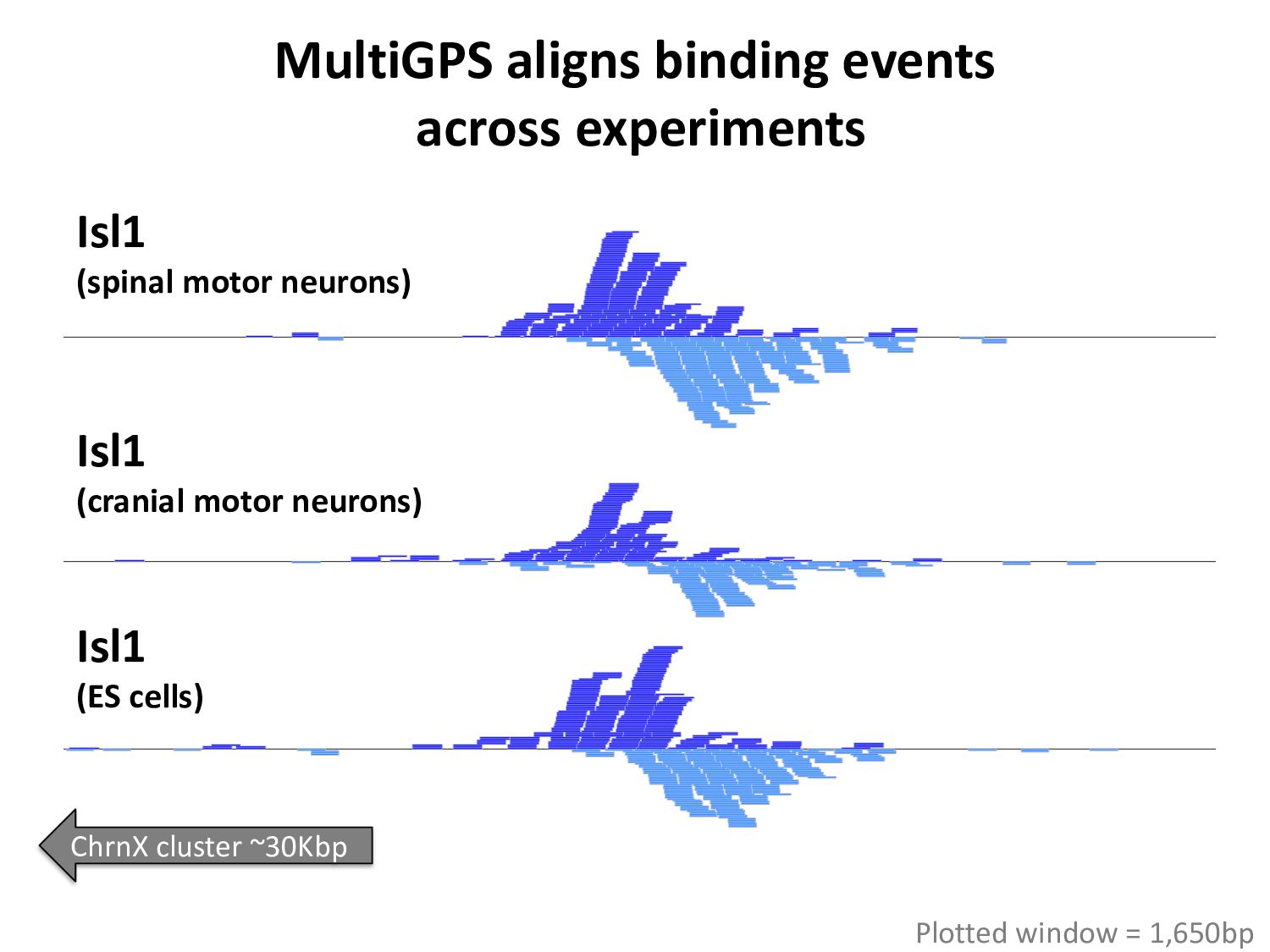
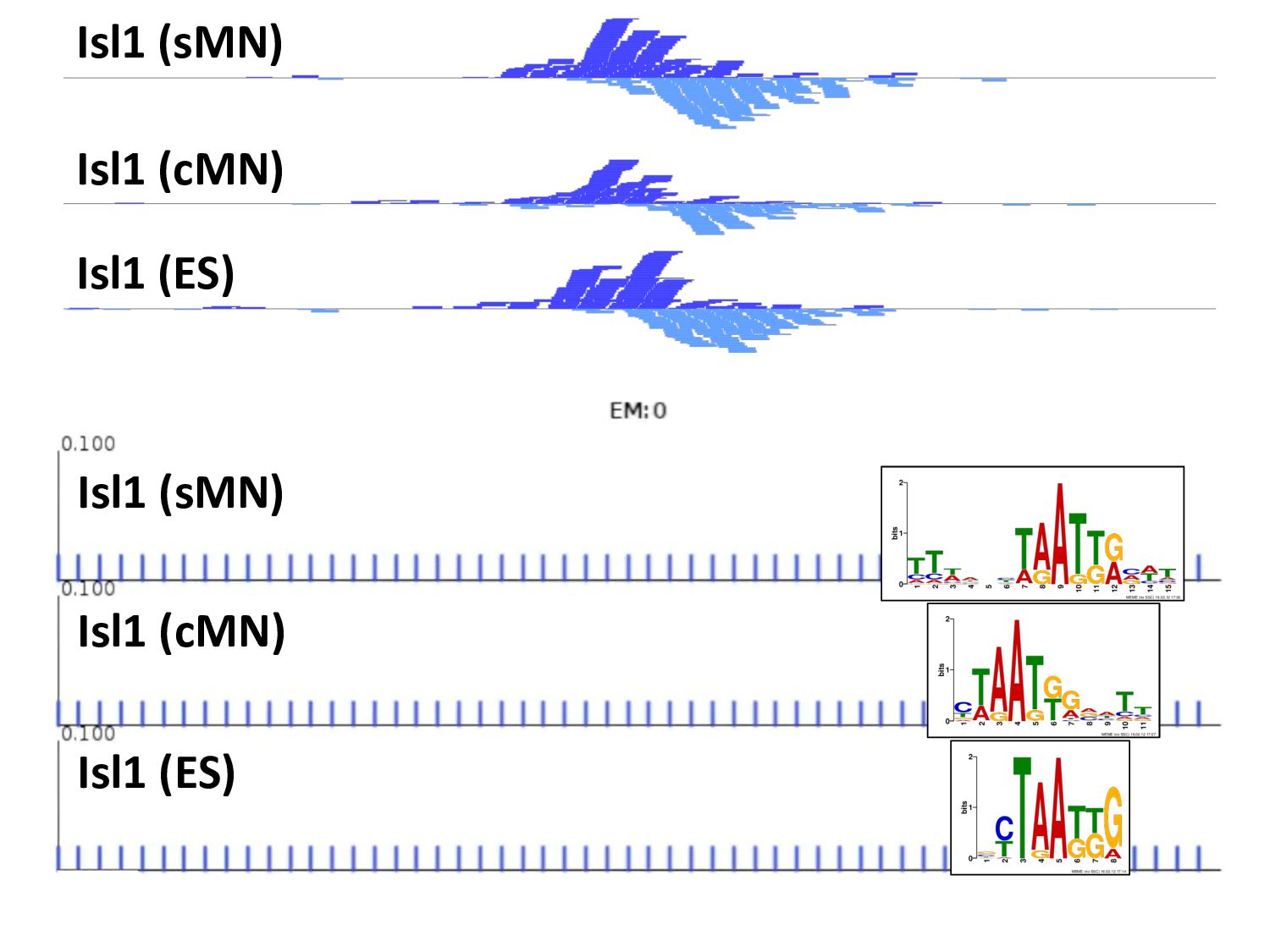
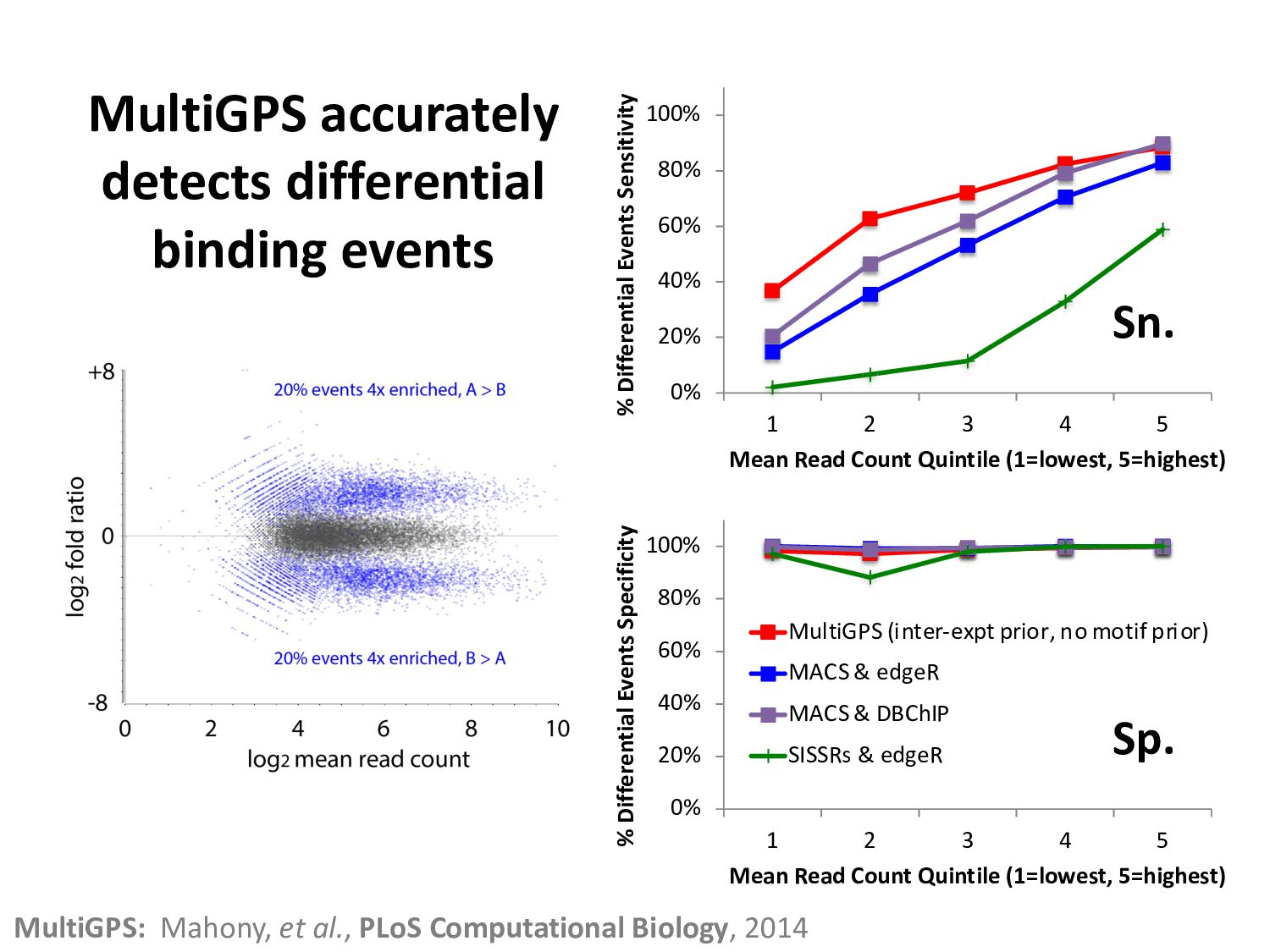
ChIP-seq peaks, or – Histone modification domains – ATAC-seq domains, or… etc. in different cell types / conditions. • Statistical significance ≠ biological significance. • How do we find loci with differential signals?
scenarios: – TF that become more highly expressed during time-course. – Histone marks after (de)methylase knock-down. • Many normalization strategies assume that most loci don’t change. • Answer - use a constant spike-in. – Spike-in exogenic material, or – Spike-in antibody against something not changing
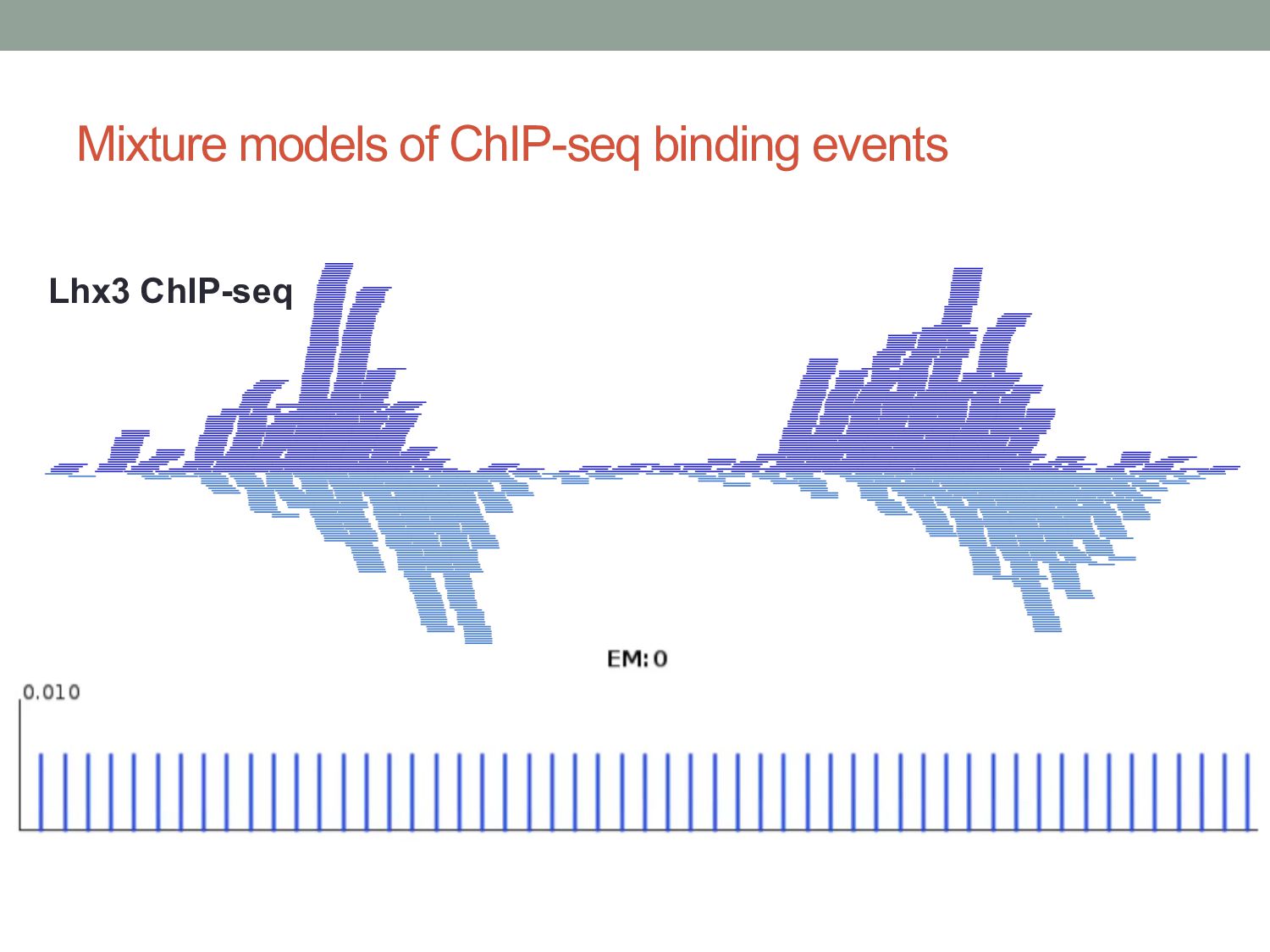
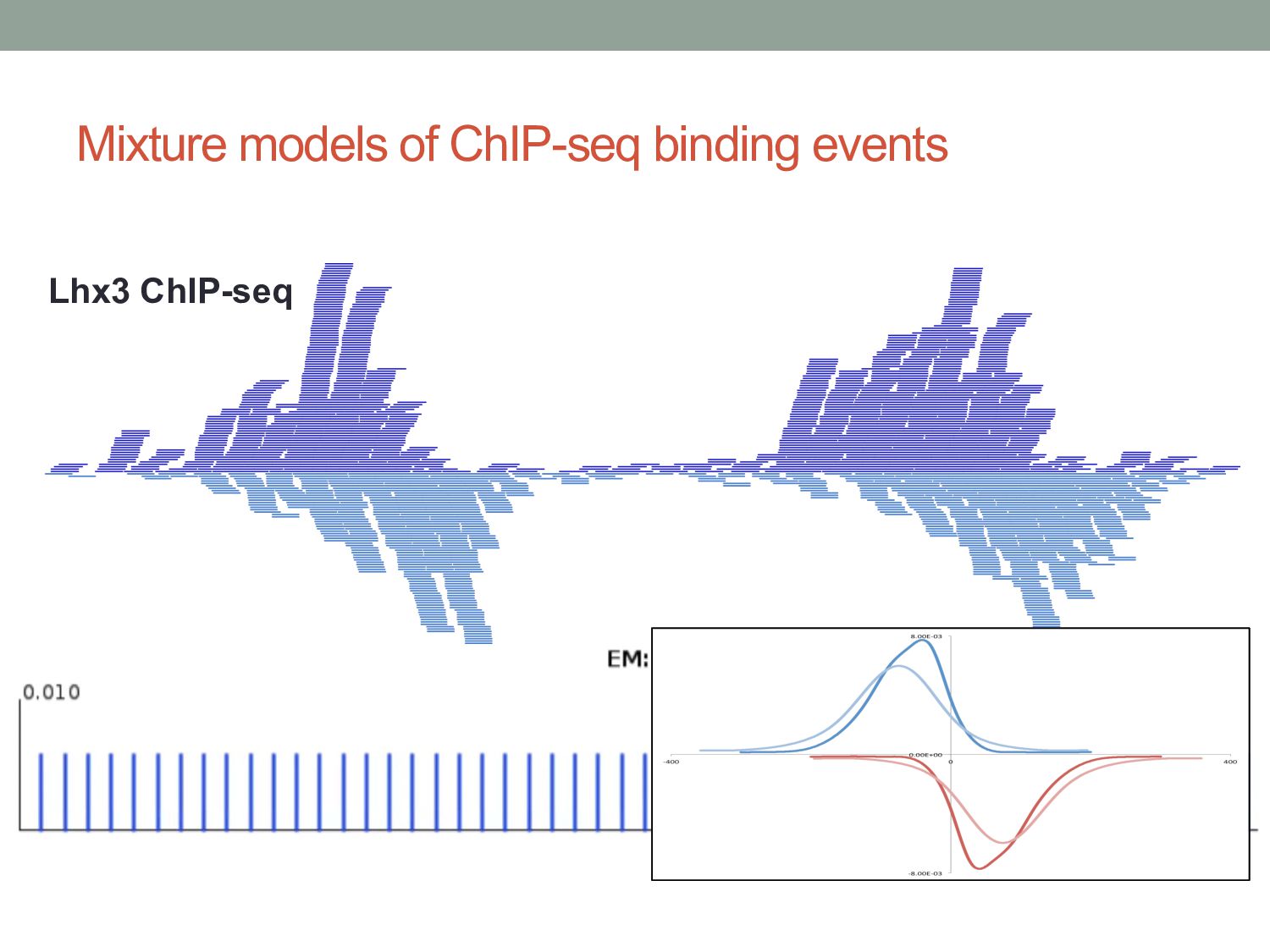
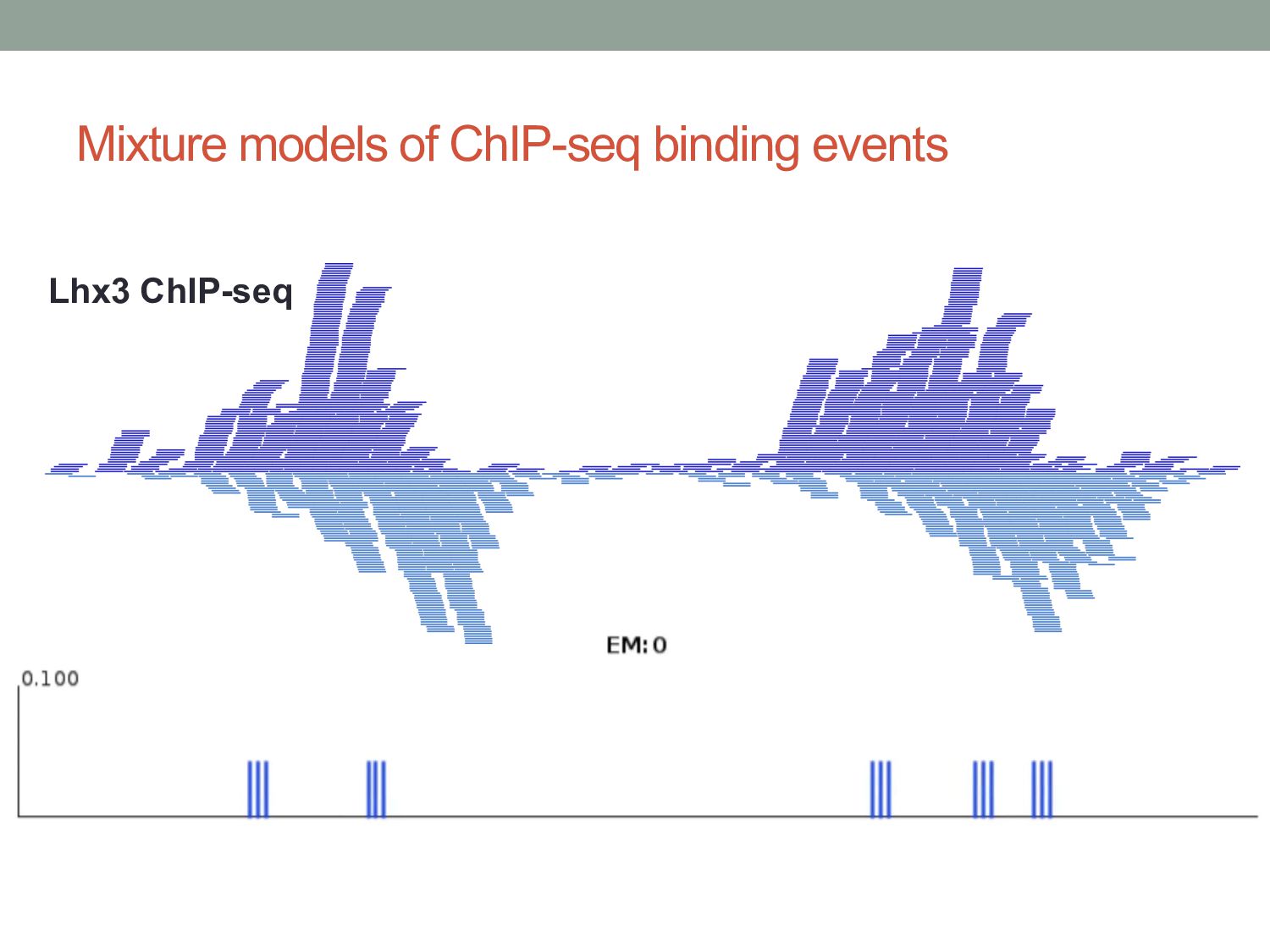
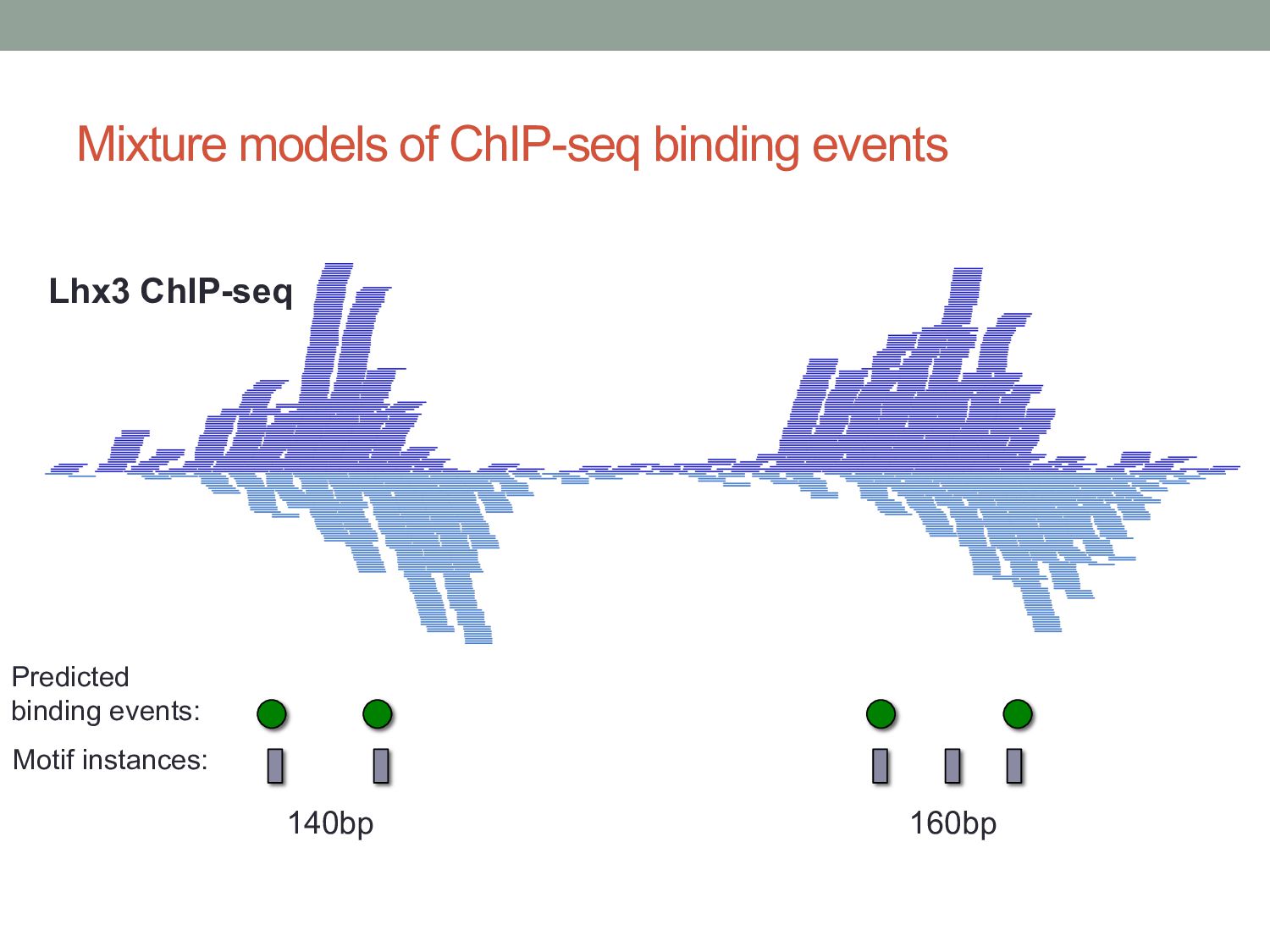
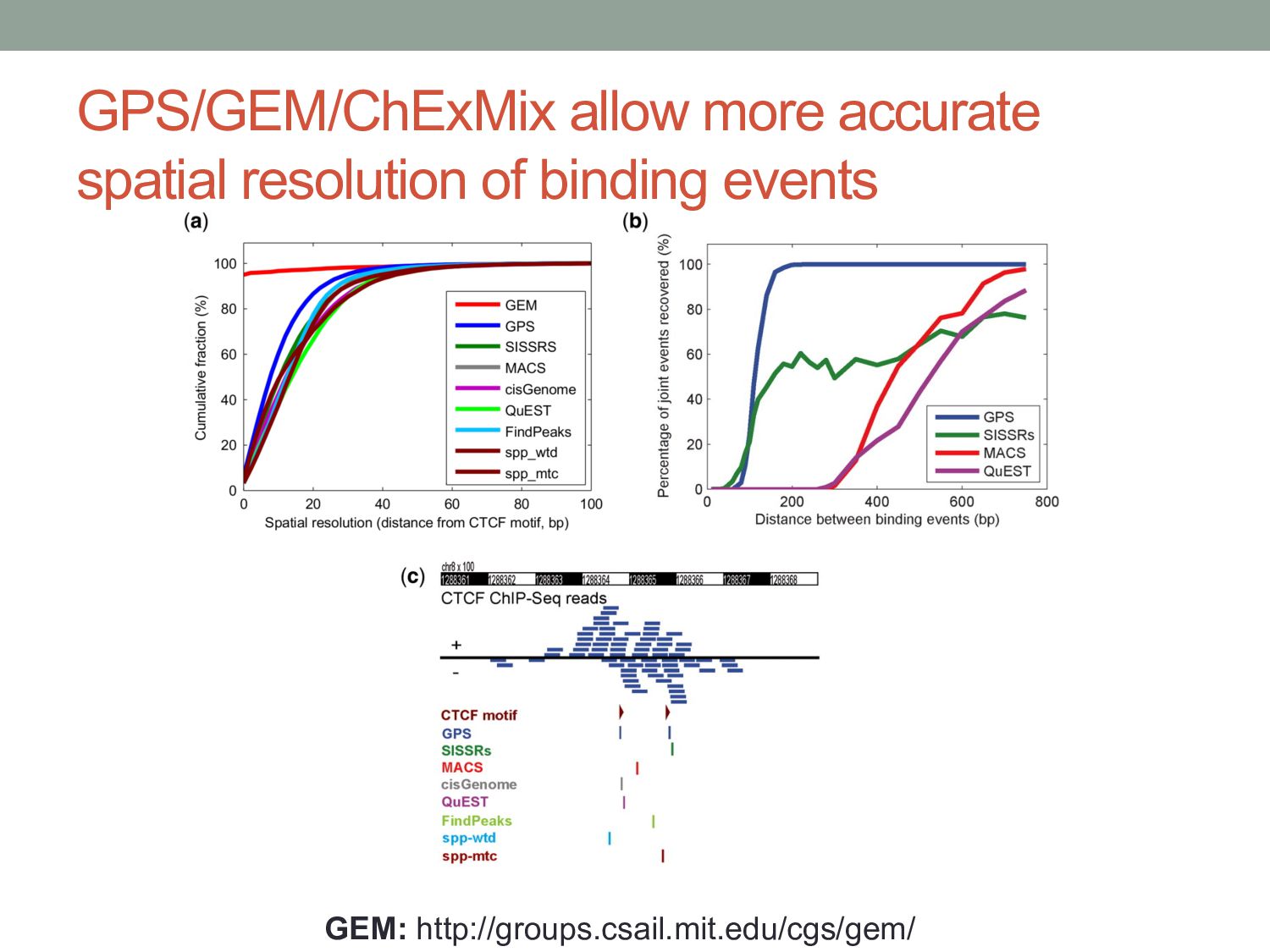
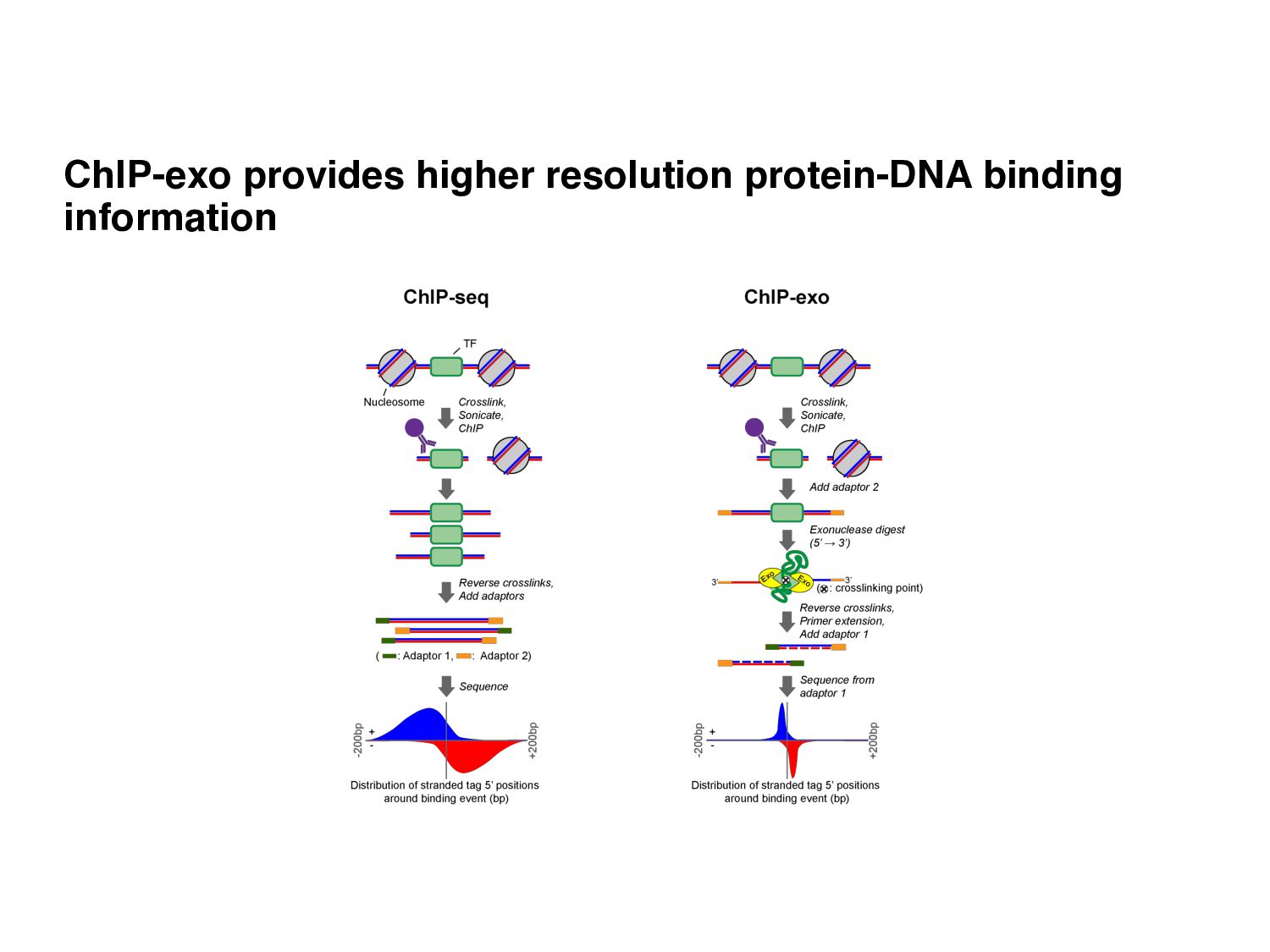
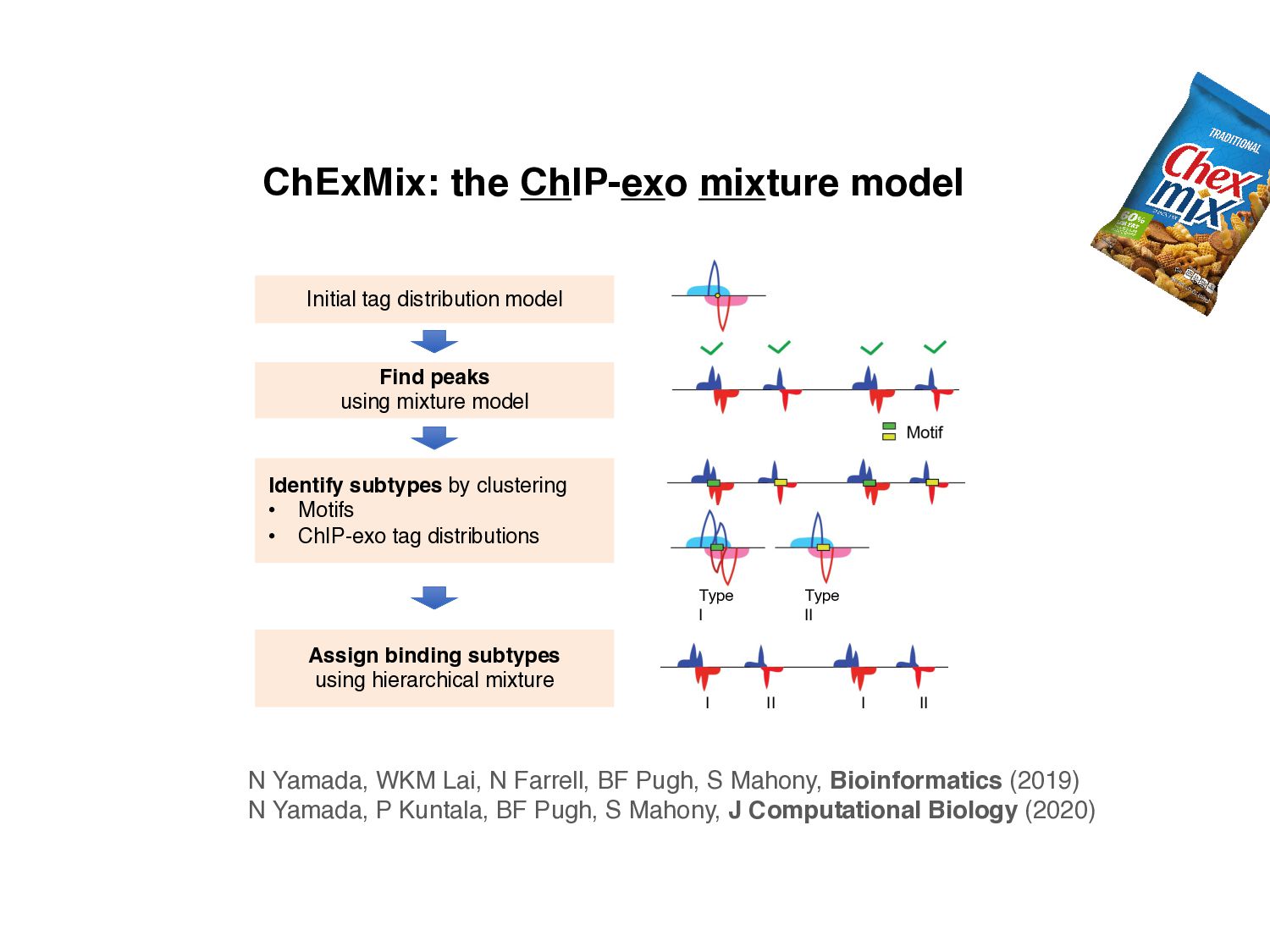
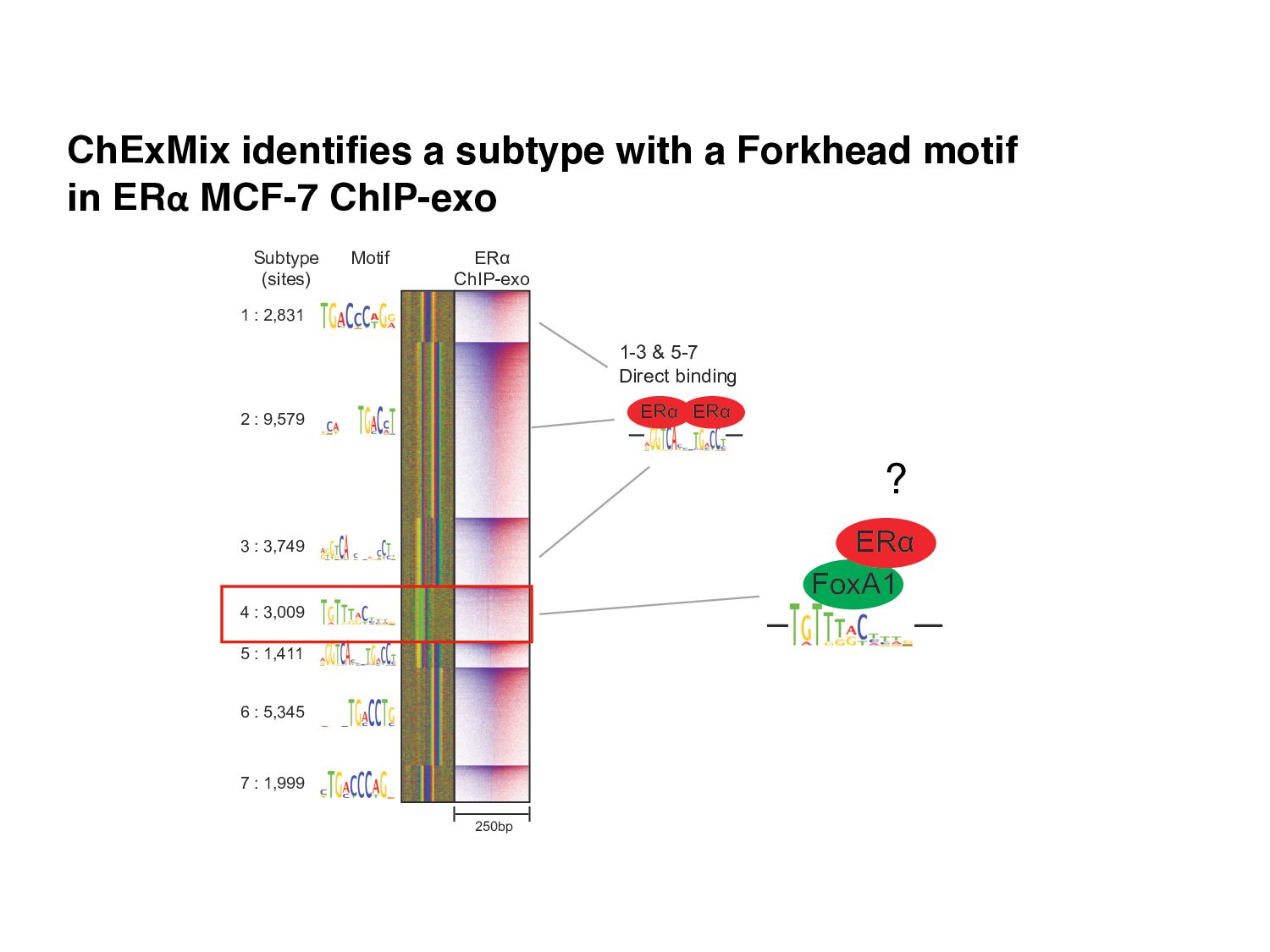
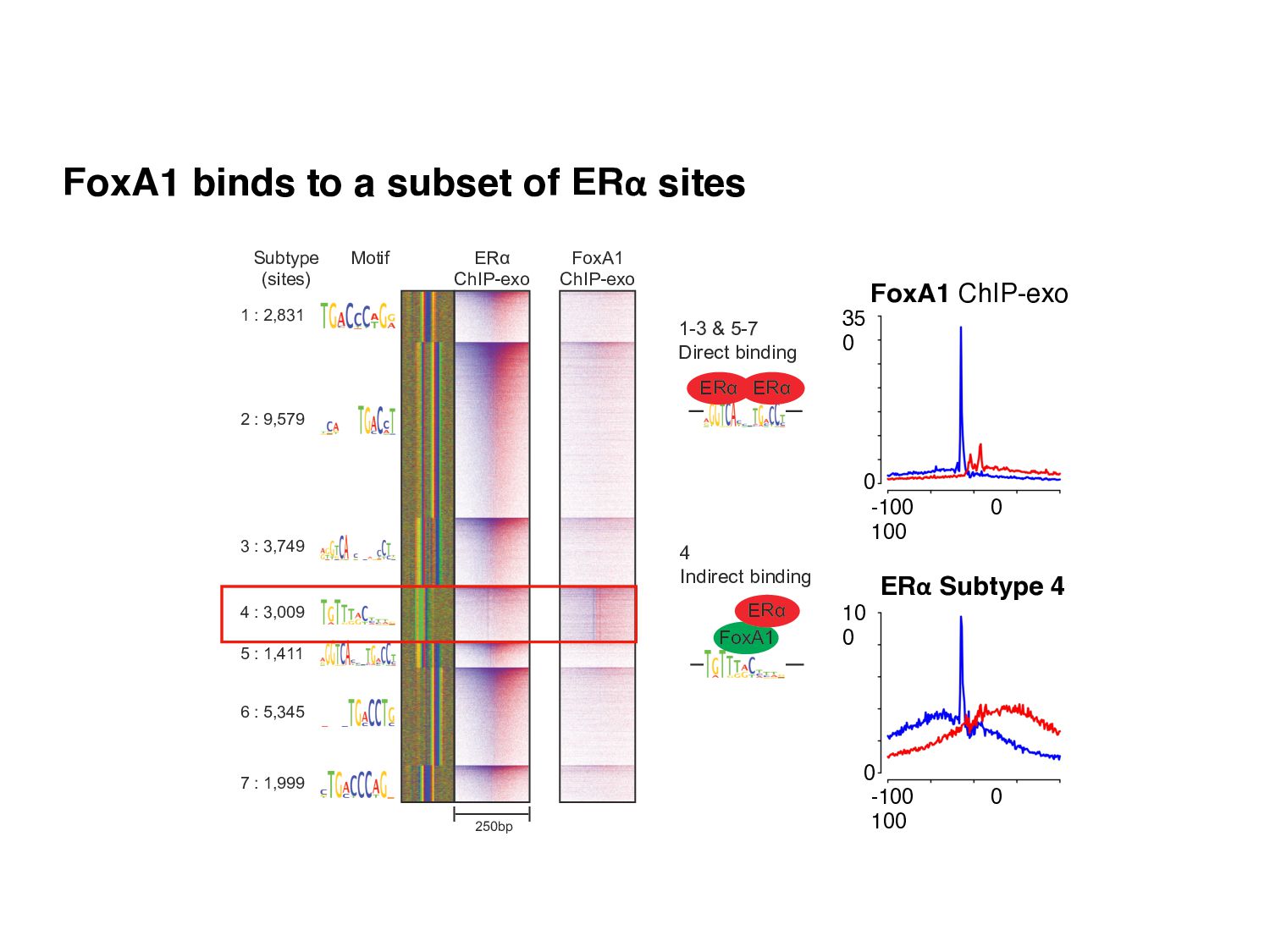
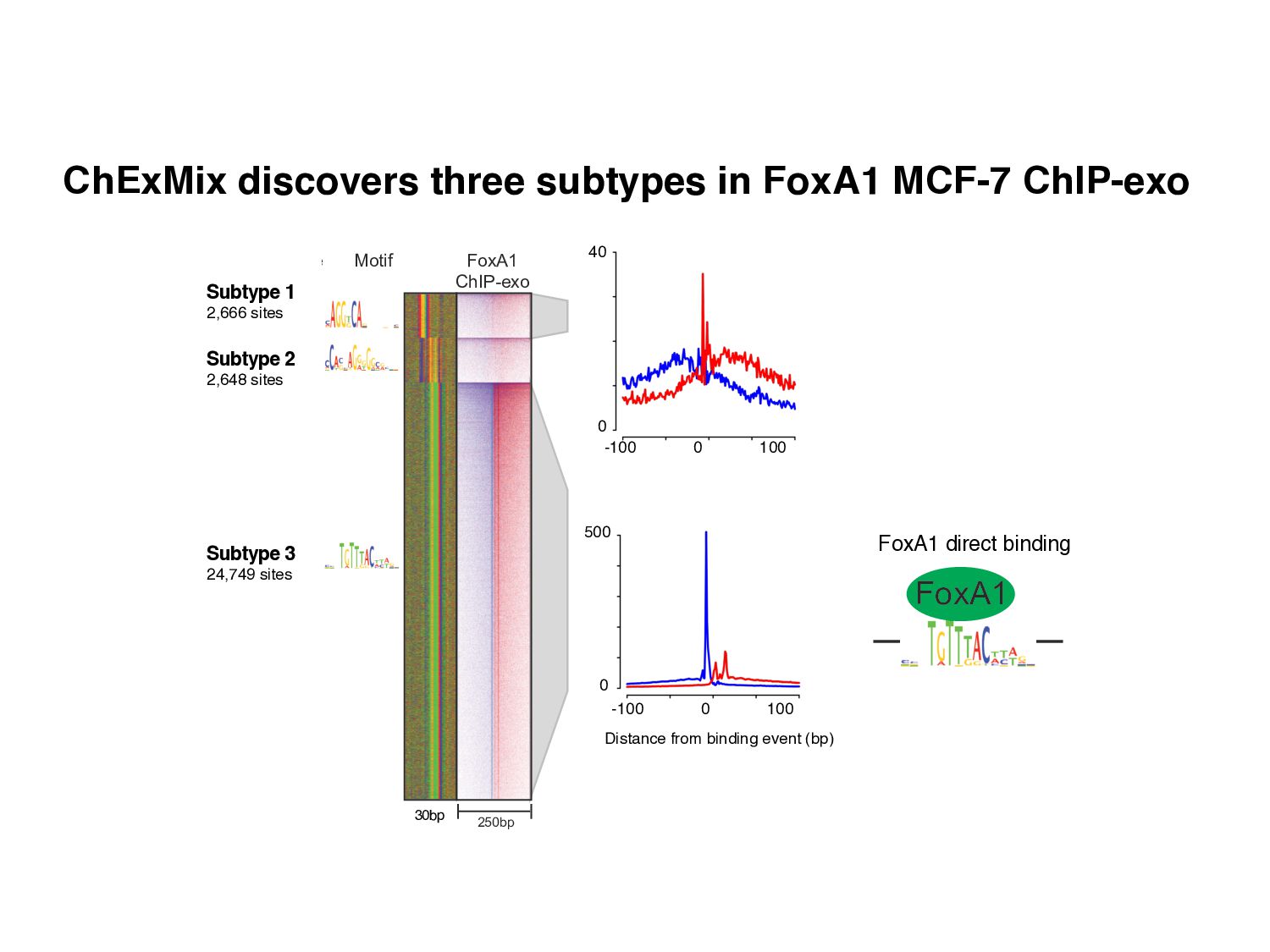
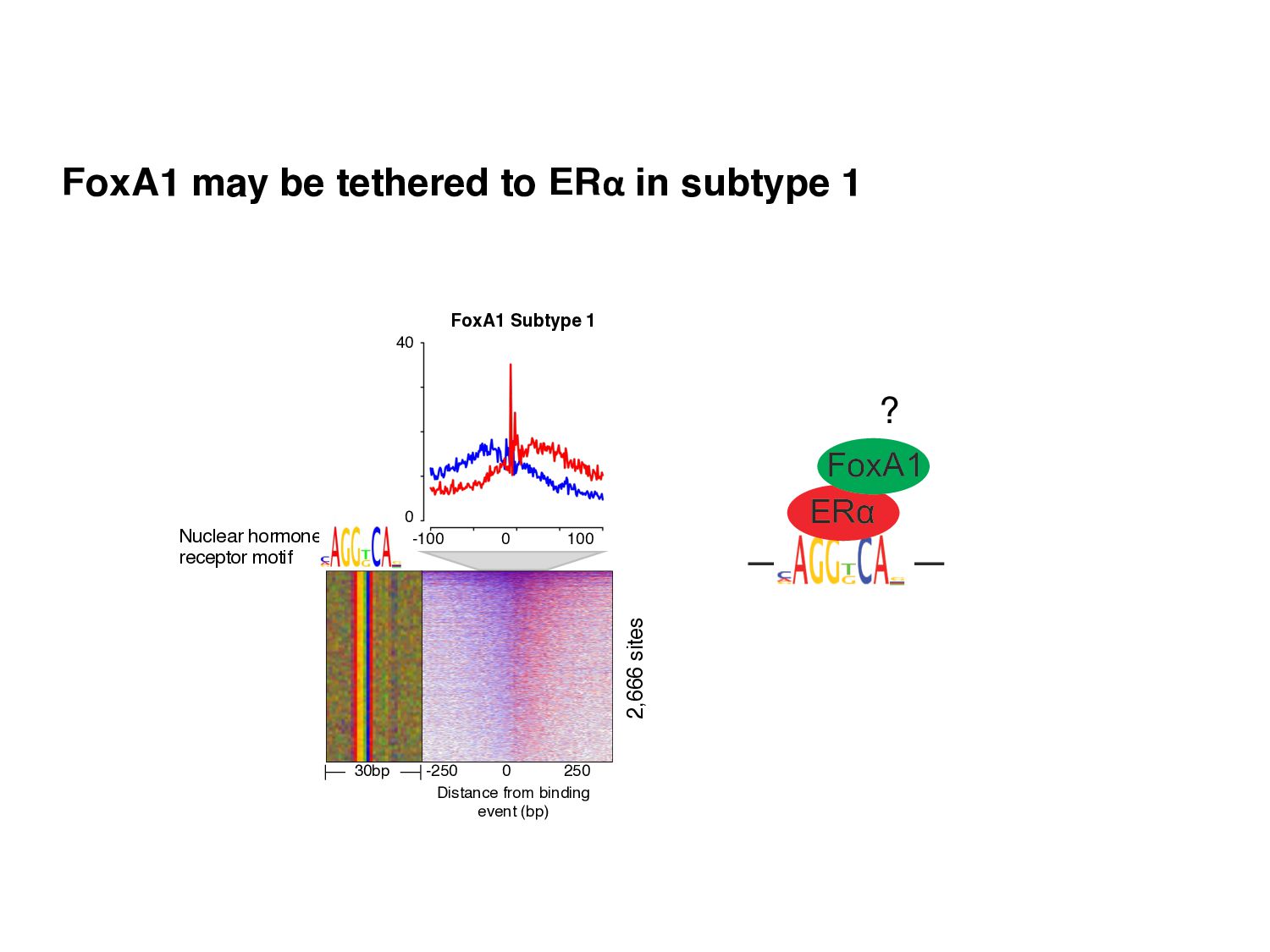
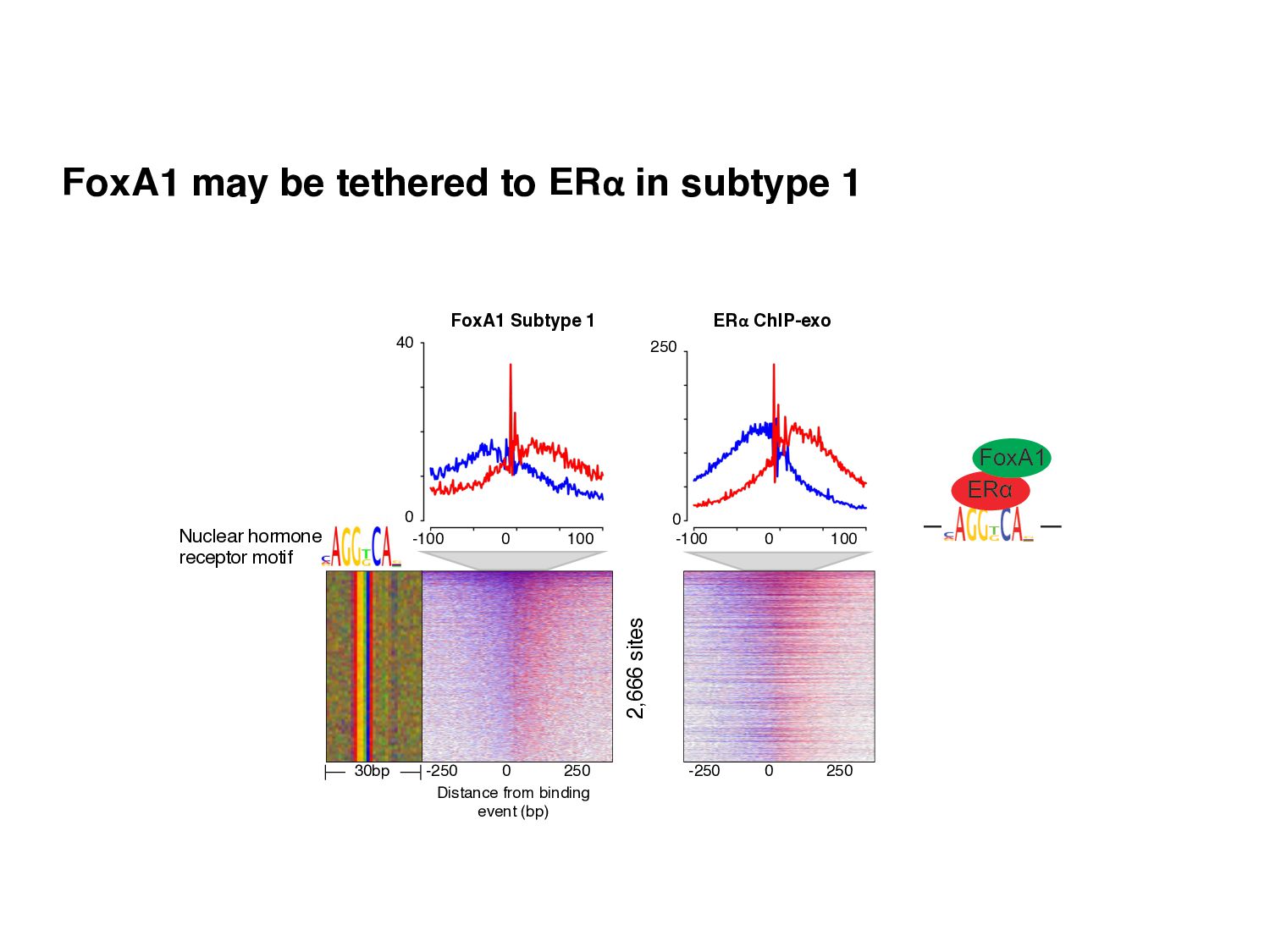
Initial tag distribution model Find peaks using mixture model Type II Type I Assign binding subtypes using hierarchical mixture II I II I ChExMix: the ChIP-exo mixture model N Yamada, WKM Lai, N Farrell, BF Pugh, S Mahony, Bioinformatics (2019) N Yamada, P Kuntala, BF Pugh, S Mahony, J Computational Biology (2020)
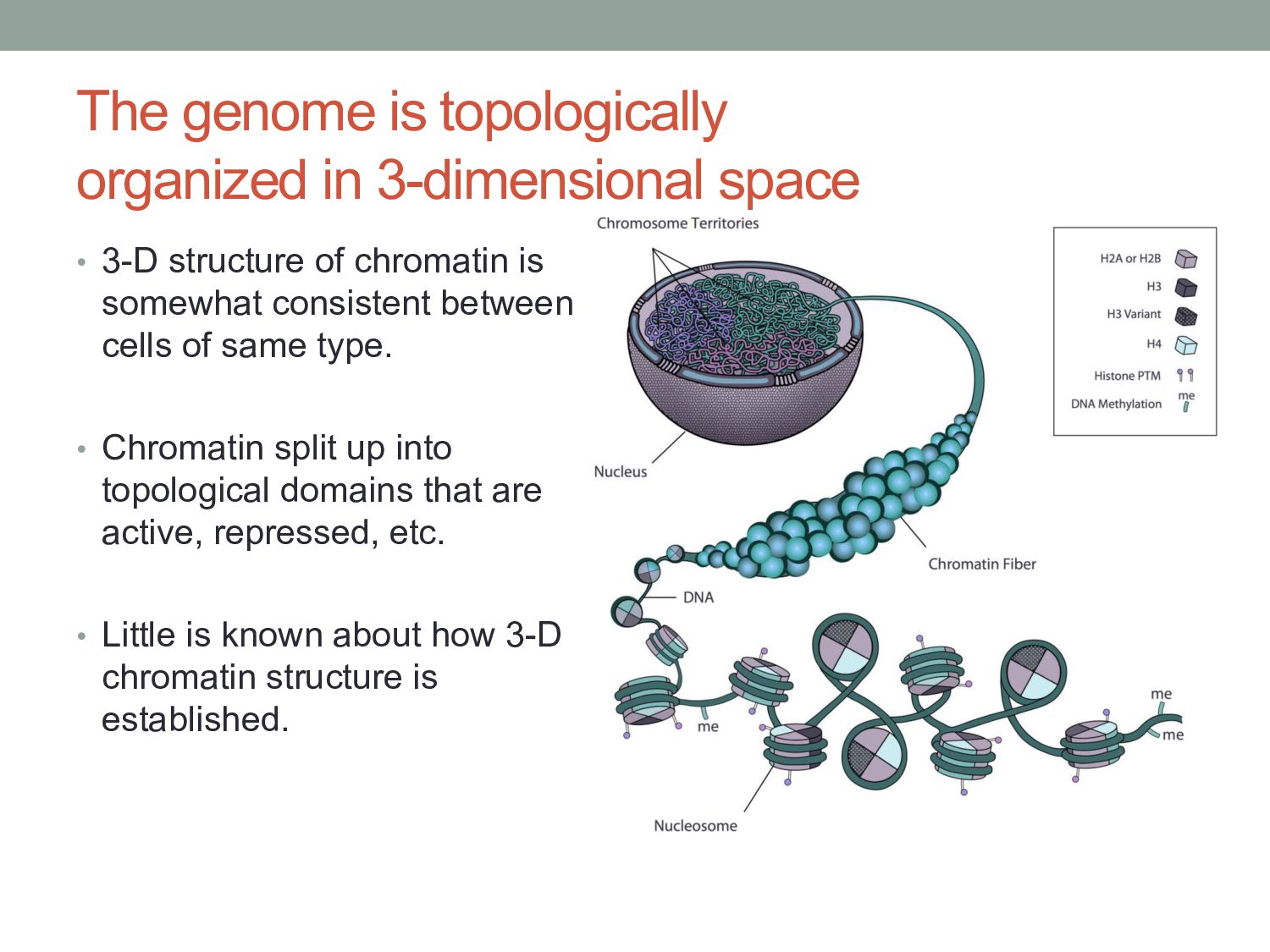
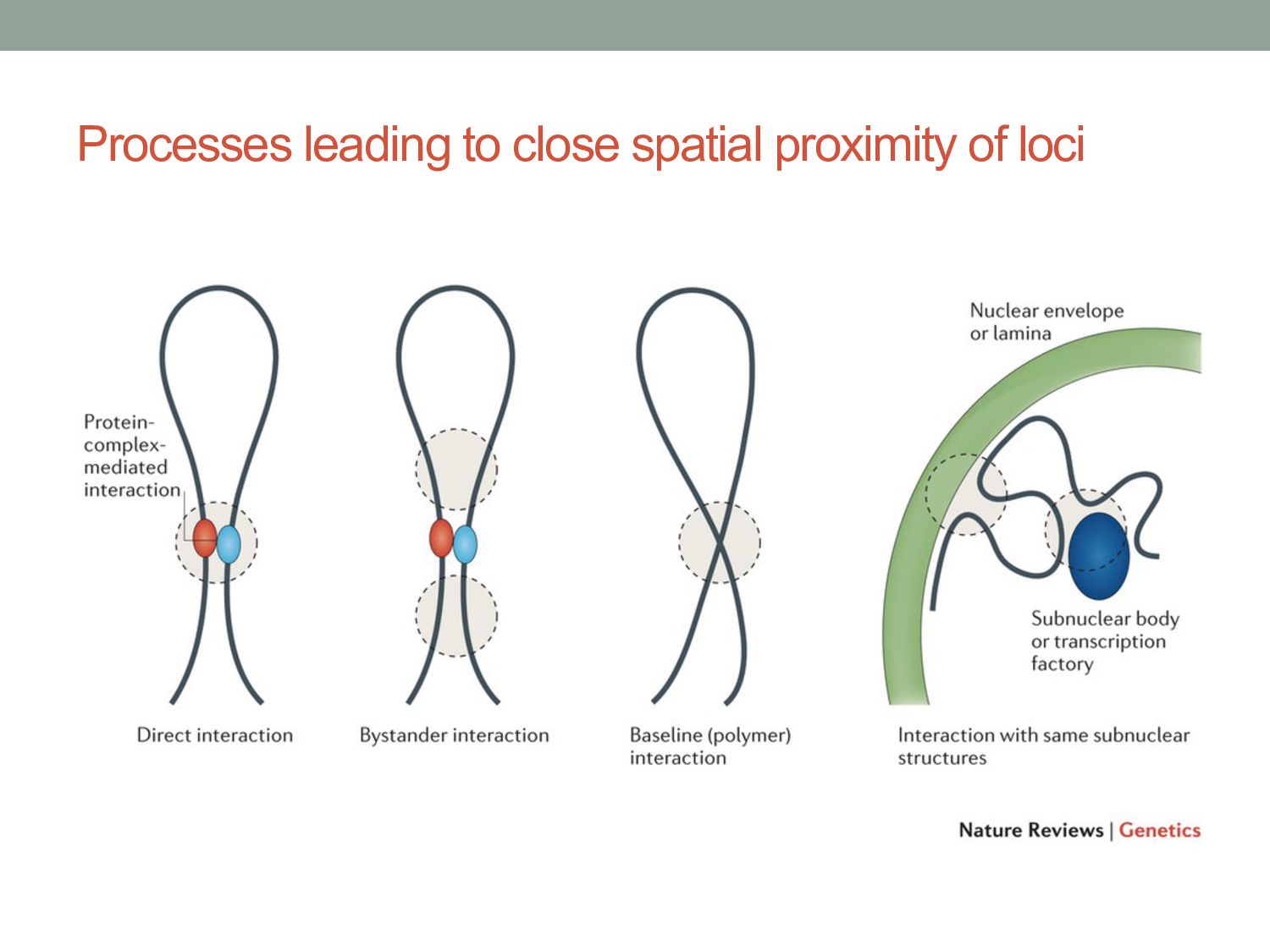
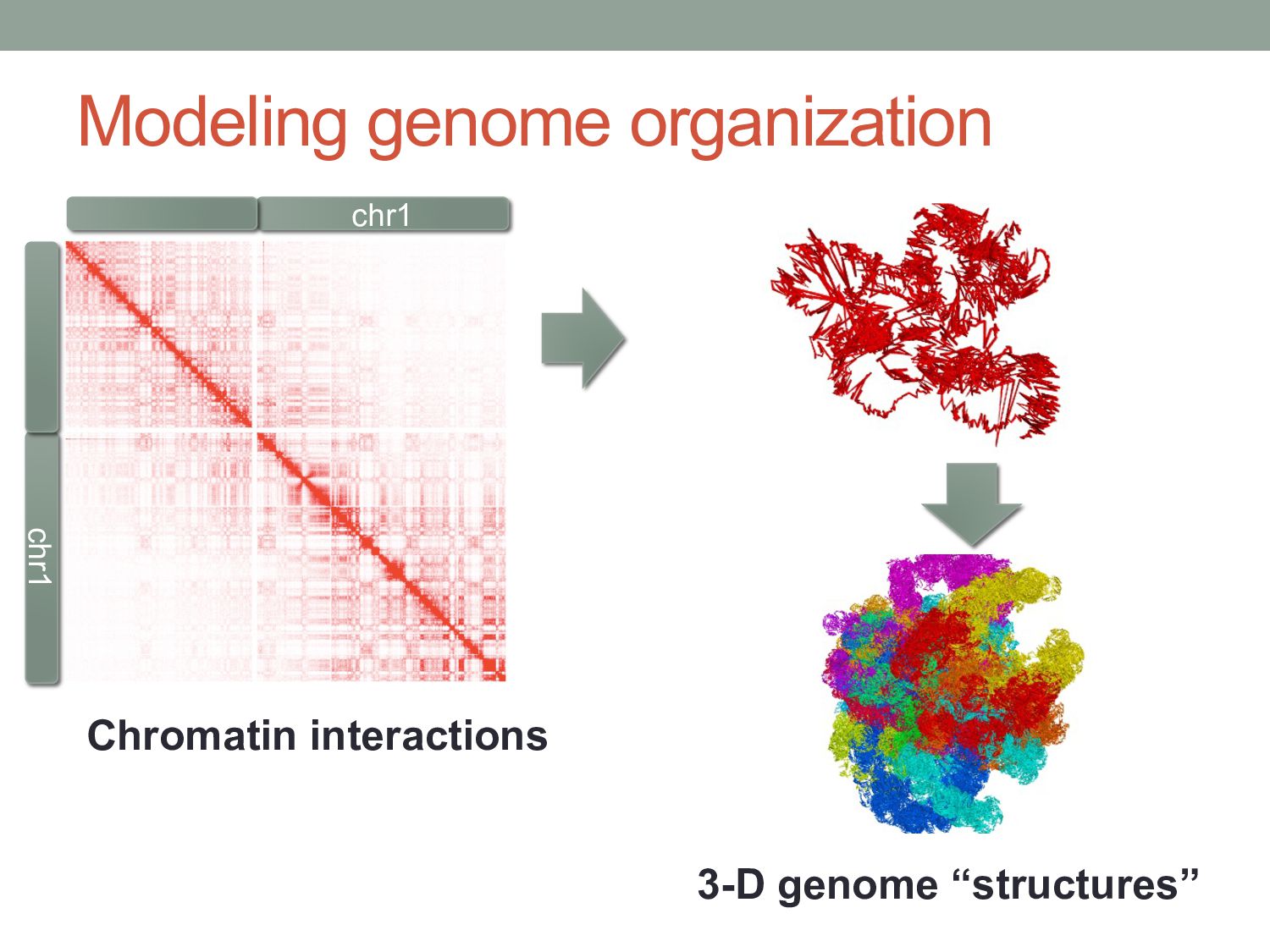
structure of chromatin is somewhat consistent between cells of same type. • Chromatin split up into topological domains that are active, repressed, etc. • Little is known about how 3-D chromatin structure is established.
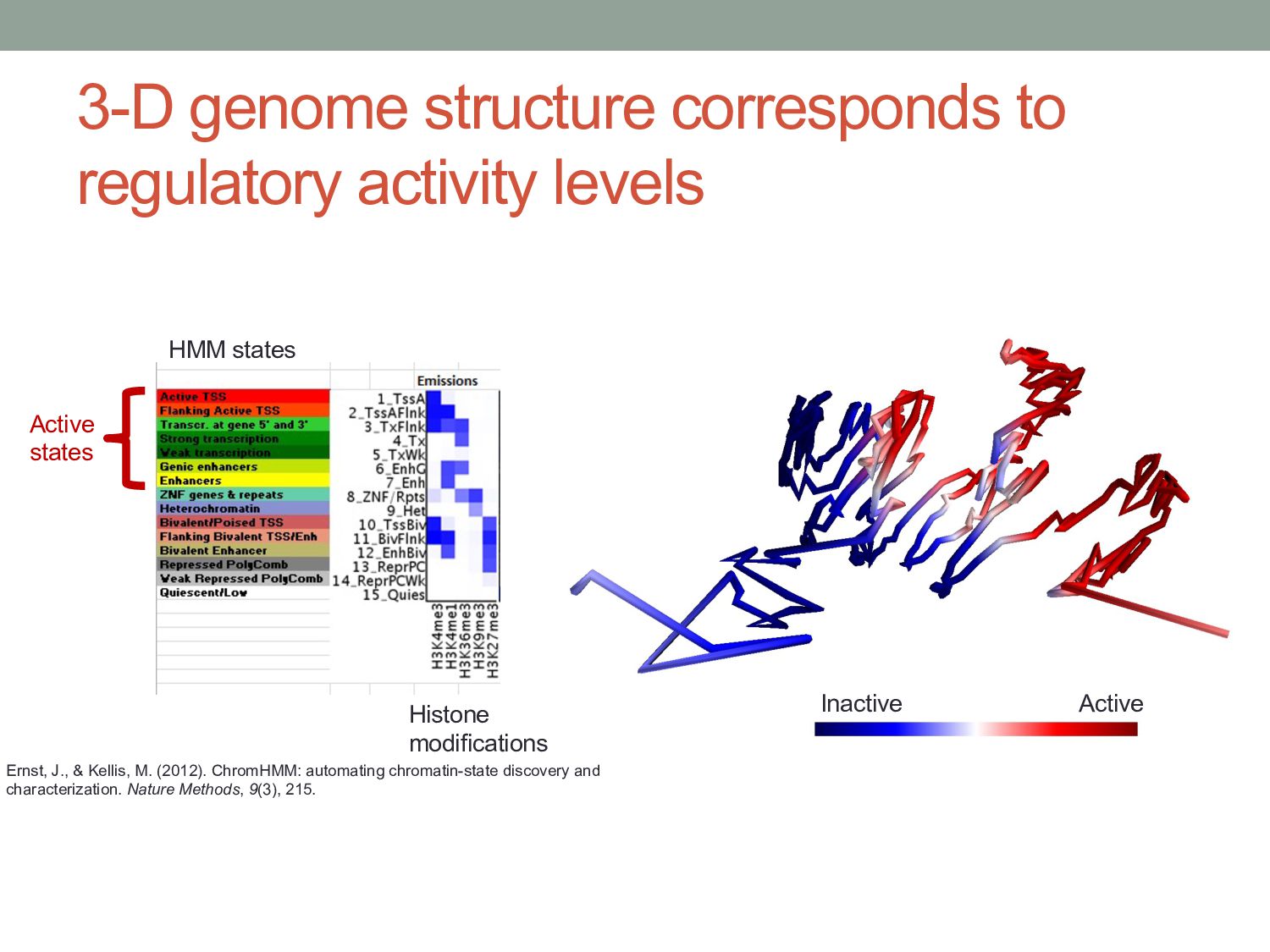
Active states HMM states Histone modifications Ernst, J., & Kellis, M. (2012). ChromHMM: automating chromatin-state discovery and characterization. Nature Methods, 9(3), 215.
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}